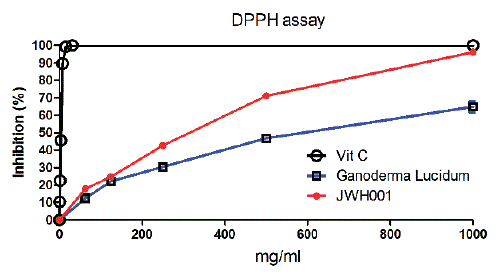
Figure 1: DPPH (1-Diphenyl-2-picrylhydrazyl) scavenging effect of H.E. mycelium and Ganoderma lucidum. Ascorbic acid (Vitamin C) was used as positive control. Data are presented as means ± SEM.

Pao-Pao Yang1,2 Chih-Yung Lin2 Tzu-Yin Lin2 Win-Chin Chiang2*
1Institute of Biotechnology and Pharmaceutical Research, National Health Research Institutes, Miaoli County, Taiwan, Republic of China*Corresponding author: Win-Chin Chiang, JOWIN BIOPHARMA Inc, Xizhi, New Taipei City, Taiwan, 9F-12, No. 97, Sec. 1, Xintai 5th Road, Xizhi Dist, New Taipei City, Taiwan (22175), E-mail: winchiang@jowinbio.com
Hericium erinaceus (H.E.) is a well-known edible and folk medicinal fungi in Japan, China and other Asian countries without harmful effects. It has been recognized that this unique mushroom is capable of keeping the brain healthy, supporting the immune system to help prevent gastric cancer and other diseases, boosting mood and concentration, decreasing inflammatory processes in the body. But more scientific researchers are needed to confirm its nutritional and medicinal effects. In the present study, we investigated the effects of Hericium erinaceus mycelium (H.E. mycelium) against 1-methyl-4-phenylpyridinium(MPP+)-induced neurotoxicity in PC12 cells and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinsonian mice. In the cell viability results, treatment with H.E.mycelium increased the cell viability in MPP+-treated cells and induced antioxidant activity in PC12 cells. H.E. mycelium also reduced MPTP-induced loss of dopamine concentration level and tyrosine hydroxylase (TH) positive cells in mice. Our results suggest that H.E.mycelium performs significant protection of dopaminergic neuron under severe conditions and is very effective in the treatment of damaged neuron in the brain to recover in the case of Parkinson’s disease.
Practical applications: In this article, we provide science-based evidence related to H.E.mycelium to be a potential effective material for the treatment and prevention of Parkinson’s disease.
Hericium erinaceus mycelium; Dopamine; Neuroprotective; Parkinson’s disease; Tyrosine hydroxylase
Parkinson’s disease (PD) is one of the most common progressive neurodegenerative disease that is characterized by the loss of dopaminergic neurons in the Substantia Nigra par compacta (SNpc) region of the brain [1], which results in motor problems including bradykinesia, akinesia, muscular rigidity, resting tremor, and postural instability [2]. In the disease model of PD, the involvement of the drug 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP), is most widely used among animal models of Parkinson’s disease [3]. The MPTP is metabolized into the toxic cation 1-methyl-4- phenylpyridinium ion (MPP+) by the enzyme monoamine oxidase B, which causes the neurotoxic effect to impair the dopaminergic nigrostriatal neurons [4].
Hericium erinaceus (H.E.) a well reputed edible mushroom, also known as monkey head mushroom in Chinese, Yamabushitake in Japanese or Lion’s mane mushroom in English, has been widely reported to use as food and folk medicine in Japan, [5] China and other Asian countries without harmful effects [6]. Several evidences demonstrated that it possesses a wide range of benefits, such as anticancer [7,8], antimicrobial [9], antioxidant [10], anti aging [11], anti-hyperglycemic [12], anti-hyperlipidemic activity [13], gastroprotective [14], immunomodulating and neuroprotective activity (Alzheimer’s Disease and Parkinson’s Disease) [15-17], protection of neuropathic pain [18], depressive symptoms [19] and presbycusis [20].
As the mycelium is inoculated in grain spawn, lion’s mane mushroom grows in large snowball-like formations, which is called the fruiting body. Hericenones, the benzyl alcohol derivatives with simple fatty acids, only exist in the fruiting body. A group of erinacines (erinacines A-K and P-S) which are diterpenoid derivatives have been identified only from the mycelium, and erinacine A is most rich in the mycelium [21]. It was demonstrated that eight of erinacines (A-I) could enhance nerve growth factor (NGF) release [21] and the erinacenes are more potent inducers of NGF synthesis than hericenones. They not only have an enhancing effect on NGF synthesis in astroglial cells in vitro but also can increase both NGF and catecholamine content in the hippocampus of rats [11]. The increased amount of NGF, in turn, enhances neuronal survival in different brain regions and substantially improves animal behavioral activity.
The aim of this present study was to explore the neuro protective effects of Hericium erinaceus mycelium (H.E. mycelium) using MPP+-treated PC12 cells or MPTP-induced PD mouse model that is associated with protection against loss of the neurotransmitter or dopaminergic neuron in vitro and in vivo.
H.E. mycelium powder (mesh size#100, RH6408) were obtained from FUNGUS BIOTECH, Co. Ltd. Yilan, Taiwan, where toxin-free and pesticide-free of Hericium erinaceus solid state fermentation was exercised and the mycelium was collected afterward and dried to the moisture content of less than 7%. The H.E. mycelium yellowish powder was then further grinded into smaller particles through a spiral jet mill (OM2 micronizer, Sturtevant Inc. Hanover, MA USA) to induce the cell wall-broken effect with a particle size distribution of D75<50 µm at FORMOSAN NANO BIOLOGY Co. Ltd, Taichung, Taiwan. The cell wall-breaking technology greatly contributed to the increased release rate of active ingredients from the fine H.E. Mycelium particle powder.
The MTS assay is a colorimetric method and usually used to assess cell proliferation, cell viability and cytotoxicity. Its protocol is based on the reduction of the MTS tetrazolium compound by viable cells to generate a colored formazan dye that is soluble in cell culture media. PC12 cell is a cell line derived from a phenochromocytoma of the rat adrenal medulla and was used in this study. The PC12cells were maintained at 3*104 cells /well in 96 wells plate with 100 µL of DMEM at 37°C in an incubator containing 5% carbon dioxide for 24 h. Briefly, after cells had attached, cells were treated with MPP+ for 72 h in the presence of ethanol extracts of test samples which was in 0.4% DMSO (Dimethyl Sulfoxide) solution. Afterward, MTS solution was added to each cell well and made it into a colored solution. The whole process was performed triplicate. The absorbance of the colored solution in each cell was measured at 570 nm using a microplate reader. To assess the neuroprotective effects of H.E. mycelium on the PC12 cells with MPP+-induced toxicity, the cells were treated with different concentration of H.E. mycelium at 8, 40, 200, 1000 µg/mL, respectively, and different concentrations of Ganoderma lucidum (Reishi) at 8, 40, 200, 1000 µg/mL, respectively, with addition of 10 mM MPP+ solution to reach the final concentration of 1 mM MPP+ in the cell. Cell viability was assessed 72 h later by measuring the absorbance of the colored solution. The survival rate of the control group was normalized as the basis for that of the other groups to compare and calculate.
The free radical scavenging activity of the tested extract was performed by using 2,2-Diphenyl-1-picrylhydrazyl (DPPH) which was at 0.1 mM Methanol solution. The extract fractions at different concentrations were prepared with distilled water/ethanol (50:50). The antioxidant standard compound, ascorbic acid, at 1 mg/mL was used as positive standard for the comparison purpose. A fresh stock solution of the standard compound was prepared before each analysis. The absorbance changes in color from deep purple to light yellow at 517 nm were measured by using a spectrophotometer after 30 min of reaction. The solution of DPPH at 0.1 mM was used as a control.
To express the radical scavenging activity, the IC50 parameter was employed and it is defined as the concentration of substrate that brings about 50% loss of the DPPH free radical. The IC50 value of H.E. mycelium is the concentration of H.E. mycelium required to inhibit 50% of the DPPH free radical. Likewise, the IC50 value of Ganoderma lucidum was so defined. The percent inhibition of DPPH free radical by a test sample calculated by using the following formula:
% of inhibition = [(dc-dt)/dc] × 100
Where dc was the absorbance of control reaction and dt was the absorbance in presence of test or standard sample.
Adult male C57BL/6 Narl mice weighing 20-30 g (8-12 weeks old) were purchased from the National Laboratory Animal Center (Taipei, Taiwan) and used in this study. Mice were kept at constant room temperature (20-22°C) and humidity (50%-70%) with a 12 h light/ dark cycle (7:00-19:00) in the Animal Centers of the National YangMing University, Taiwan. Standard diet and water were available Ad libitum during the experiment. The care of animals was carried out in accordance with institutional and international standards (Principles of Laboratory Animal Care, NIH); and all experiments were approved by the Institutional Animal Care and Use Committees of the National Yang-Ming University, Taiwan (IACUC No. 1050306). All studies involving animals were conducted in accordance with the ARRIVE guidelines [22,23].
Five groups (fifteen mice in each group) were randomly assigned to a control group (saline, i.p.) plus H2O (p.o.), MPTP group (20 mg/ kg, i.p; Tokyo chemical Industry, TCI) plus H2O (p.o.) and MPTP (20 mg/kg, i.p.) plus H.E. mycelium groups (0.1 g/kg, 0.3 g/kg and 1 g/kg., p.o.). Mice were received intraperitoneal injection of saline or MPTP once a day in the beginning 5 consecutive days and oral administration of H2O or H.E. mycelium in the duration of 30-day period. The control group of animal received an equivalent volume of saline. The animals were sacrificed 30 days after the last treatment of five groups and then brains were dissected to the left and right cerebrum. The right cerebrum was removed to determine immunohistochemistry for tyrosine hydroxylase and the left cerebrum of striatum was rapidly removed to determine concentration of dopamine neurotransmitter.
The mice were sacrificed under pentobarbital after completion of the treatment. The left cerebrum of striatum was quickly removed and homogenized in a stock solution containing 0.1 M HClO4, 0.1 mM EDTA, 0.1 mM Na2S2O5 and centrifuged at 13,000 rpm for 10 min at 4°C. The supernatant was harvested, filtered with 0.45 µm poresize filters, and reserved at -80°C until analysis. The concentration of dopamine was determined by using High-Performance Liquid Chromatography (HPLC) with electrochemical detection.
HPLC electrochemical detection was performed to quantify the concentration of dopamine. The HPLC system consisted of a pump (BAS PM-92E; Bioanalytical Systems, West Lafayette, IN, USA), a refrigerated microsampler (CMA/200) and a sample injector (CMA/240) with a 20 µL loop (CMA, Stockholm, Sweden), and a digital amperometric electrochemical detector (Decade II; Antec Leyden BV, Zoeterwoude, The Netherlands). For the determination of the dopamine concentration, 15 µL samples were injected into the HPLC system. The mobile phase consisted of 0.74 mM sodium1-Octanesulfaoate (SOS), 100 mM phosphate sodium salt, 0.027 mM EDTA, 2 mMKCl, 125 mL methanol, which was delivered at a flow rate of 500 µL/min. A reversed-phase C-18 column (100 × 4.6 mm, 2.6 µm) was used for sample separation. The applied potential of the glassy carbon electrode was 650 mV to the reference electrode (Ag/AgCl), and the range setting was 5 nA for the determination of neurotransmitters. The data acquisition and analysis were performed using EZChrom software (Scientific Software, San Ramon, CA, USA). .
For measuring tyrosine hydroxylase immunohistochemistry, the mice were anesthetized with pentobarbital after completion of the treatment. The right cerebrum was removed and soaked at 4°C in the paraformaldehyde (PFA) alone, and maintained in 30% sucrose at 4°C until they sank. The cerebrum was sectioned at 10 µm. All sections were stained for tyrosine hydroxylase measurements.
Data were expressed as the mean ± SEM. Analysis of variance was used to access the statistical significance for repeated measures of the data, and the differences among the individual mean values in different groups were analyzed by ANOVA followed by the Newman-Keuls test. The differences were considered to be significant at p<0.05.
The DPPH radical scavenging assay was determined spectrophotometrically. The radical scavenging capacities of the tested extract were performed by DPPH assay and results were shown in figure 1. DPPH assay was one of the most widely and commonly used methods for screening antioxidant activity of plant or mushroom extracts. In the figure 1, DPPH scavenging activities of H.E. mycelium, and reference antioxidants capacities of Ganoderma lucidum (Reishi) obtained from FUNGUS BIOTECH, Co. Ltd., Yilan, Taiwan and ascorbic acid were analyzed in this study. The concentrations of H.E. mycelium at the range of 62.5 and 1000 µg/mL exhibited positive DPPH scavenging activities in a concentration-dependent manner as had shown in figure 1. The higher the concentration of H.E. mycelium at, the more the DPPH radical scavenging activity. As compared, Ganoderma lucidum showed a relatively lower radical scavenging effect than H.E. mycelium. The radical scavenging effect of the H.E. mycelium can also be observed by comparing IC50 value. A lower IC50 value indicates a higher antioxidant activity. The IC50 value of the H.E. mycelium was 217.2 µg/mL, compared to that of Ganoderma lucidum with 550.4 µg/mL. It indicated H.E. mycelium had a better radical scavenging effect than Ganoderma lucidum. Overall, these findings support that H.E. mycelium exerted a good antioxidant activity in PC12 cells.
Figure 1: DPPH (1-Diphenyl-2-picrylhydrazyl) scavenging effect of H.E. mycelium and Ganoderma lucidum. Ascorbic acid (Vitamin C) was used as positive control. Data are presented as means ± SEM.
The protective ability of H.E. mycelium from the cytotoxicity of MPP+ in PC12 cells was measured by using the MTS assay. The results of the measurement, as shown in figure 2, revealed the decrease of cell viability of PC12 cells after exposure to 1 mM MPP+ for 72 h. In the presence of 1 mM MPP+, H.E. mycelium at the concentration range of 40, 200 and 1000 µg/mL showed a statistically significant protective effect in a concentration dependent manner. A high concentration exerted a better neuroprotective effect. But, Ganoderma lucidum showed a significant protective effect in the PC12 cells only at high concentration of 1000 µg/mL, as shown in figure 2. It can be concluded that H.E. mycelium exerted a very good protective effect against the MPP+ neurotoxicity in PC 12 cells.
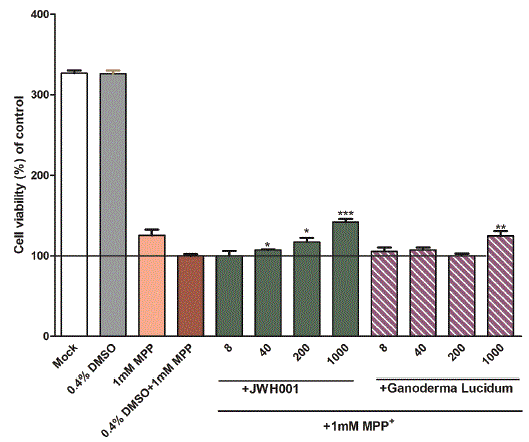
Figure 2: H.E. mycelium protected MPP+ induced neurotoxicity in PC12 cells. PC12 cells were treated with H.E. mycelium or Ganoderma lucidum with MPP+. Cell viability was measured by MTS assay. Data are presented as means ± SEM. One way ANOVA and Newman-Keuls tests were used to analyze the data. *P<0.05; **p<0.01; ***p<0.001 compared to 0.4% DMSO with 1 mM MPP+.
The protective effect of H.E. mycelium against MPTP-induced toxic damage on dopaminergic neuron can be interpreted by looking at the reduction of decreased dopamine level in the striatum of the brain after H.E. mycelium treatment. The dopamine concentration was measured by using HPLC. From figure 3, one can recognize that five injections of MPTP (20 mg/kg, i.p.) reduced to about 46% of dopamine level of striatum in mice, compared to the control group. As expected, the oral administration of H.E. mycelium at 0.3 and 1 g/kg increased the dopamine level to 56% and 79%, respectively, which indicated that the protective effect of H.E. mycelium was 20% and 70% at 0.3 and 1.0 g/ kg, respectively, as compared to the MPTP group [24]. These results demonstrated that H.E. mycelium exerted the protective effect against the MPTP-induced dopamine neuron damage.
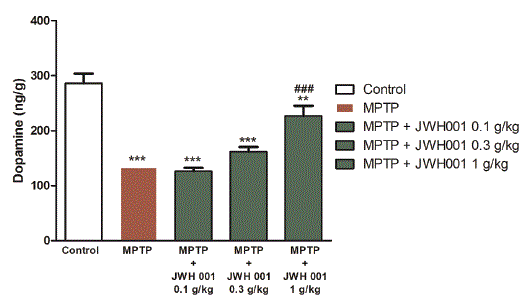
Figure 3: H.E. mycelium prevented MPTP-induced level of dopamine concentration. H.E. mycelium (0.1, 0.3 and 1 g/kg) and MPTP on the concentration of dopamine level in striatum. Data are presented as means ± SEM (n=10-15). One way ANOVA and Newman-Keuls tests were used to analyze the data. **p<0.01; ***p<0.001 compared to when compared with control; ##P<0.01 when compared to MPTP.
The enzyme tyrosine hydroxylase (TH) converts the amino acid L-tyrosine into 3,4-dihydroxyphenylalanine (L-DOPA) which then converts to dopamine by decarboxylation and then norepinephrine and epinephrine are produced in series in the pathway in both central and peripheral nervous systems [25]. The biosynthetic of dopamine pathway is illustrated in figure 4. Therefore, the activity of tyrosine hydroxylase can influence the dopamine level of striatum in brain tissue and the reduction of tyrosine hydroxylase activity represents the damage of dopaminergic neurons in the brain. Figure 5 shows a large number of TH-positive cells in the control group and MPTP caused 63% reduction in TH-positive cells in mice compared with the control group. After treatment of H.E. mycelium at 0.1, 0.3 and 1 g/kg in the MPTP-induced mice, it improved the TH density in the substantia nigra area of the brain by 18%, 78% and 100% at these three concentrations respectively, compared with the MPTP damage group. These findings confirmed that H.E. mycelium exerted protection from MPTP-induced death of TH-positive neurons in corpus striatum of mice.
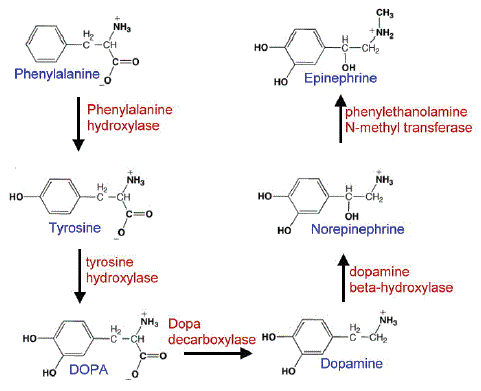
Figure 4: The biosynthetic pathway of dopamine neurotransmitters. Tyrosine hydroxylase is the rate limiting enzyme of the pathway. Phenylalanine hydroxylase converts phenylalanine to tyrosine, tyrosine hydroxylase hydroxylates tyrosine to L-DOPA. DOPA is converted to dopamine by decarboxylase. Dopamine converts to Norepinephrine by dopamine-β-hydroxylase and norepinephrine to epinephrine by phenylethanolamine N- methyl transferase.
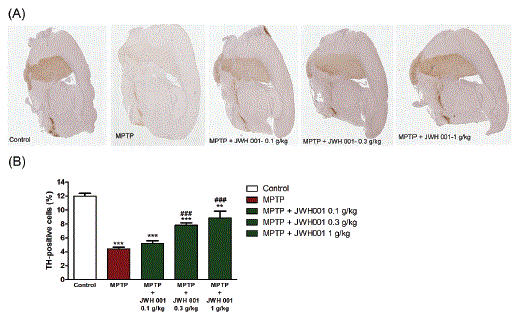
Figure 5: H.E. mycelium prevented MPTP-induced death of TH (Tyrosine Hydroxylase)-positive neuron in striatum in mice.
(A). TH expression in striatum was assessed by immunohistochemistry.
(B). Quantitative analysis of TH-positive cells in the striatum of mouse brain. Dopamine neurons from mice are resistant to MPTP neurotoxicity. There is significant difference in the average number of TH positive neurons between the striatum of 0.3, 1 g/kg and that of MPTP group. Data are presented as means ± SEM (n=10-15). One way ANOVA and Newman-Keuls tests were used to analyze the data. **P<0.01; ***p<0.001 when compared to control; ###P<0.001 when compared to MPTP.
In this present study we demonstrates that the neuroprotective effect of H.E. mycelium against MPP+-induced toxicity in neuronal PC12 cell lines and MPTP-induced striatal dopamine neuron damage in mice model. Our data strongly suggests that H.E. mycelium intake could be a potential treatment of PD (Parkinson’s disease).
Parkinson’s disease (PD) is characterized by a progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc) region of the brain [1], and N-methyl-4-phenyl1,2,3,6-tetrahydropyridine (MPTP) has been widely used to induce PD in the animal model. MPTP induces PD through the death of dopaminergic cell by its active metabolite, 1-methyl-4-phenyl-2,3,- dihydropyridinium (MPP+) [3]. By using MPP+ in PC12 cells, as a cellular model of PD, we have elucidated the role of H.E. mycelium in modulating and reducing the loss of dopaminergic neurons. In the PC12 cell viability experiment, H.E. mycelium exhibited positive protective effect to dopamine neuron in mice which represents the similar neuroprotective properties in dopamine neurons to human PD [26,27]. Numerous studies have shown that MPP+-induces neurotoxicity in cell and zebrafish [28], and induces loss of dopaminergic neurons [29- 31]. Administration of MPP+ (1 mM) induced cell death in PC12 cells is in consistent with several previous studies. MPP+-induced loss of dopaminergic neurons indicates the selective damage to dopamine neurons in cells. H. erinaceus extract promoted NGF synthesis by hericenones from fruit bodies (Hericenone C-H) and by erinacines from mycelium (erinacines A-I) [15]. H.E. mycelium reduced MPP+- induced PC12 cell toxicity in a concentration dependent manner, indicating that H.E. mycelium could contribute to the protective effect of dopaminergic neuron in PC12 cells. In 2002, Park YS, et al. [32] reported that H. E. enhanced the synthesis of NGF (Nerve Growth Factor) and BDNF (Brain-Derived Nerve Factor) in PC12 cells. BDNF and NGF can be strongly expressed by dopaminergic neurons in SNpc. But, in human and animal models of Parkinson’s disease, the expression of NGF and BDNF levels are decreased [33,34]. Thus, NGF level corresponds very well to the severity of PD. By summing up all the evidence, a theory can be formed, in which erinacines of H.E. mycelium upon ingested are transported to the brain where significant amount of NGF molecules are formed. The NGF molecules then promote brain nerves in substantia nigra to grow where dopaminergic neuron produces dopamine to a level that alleviates the Parkinson’s disease.
In terms of effective dosage of H.E. mycelium, it was shown in this study that it performed much better than Ganoderma lucidum. Although it has been reported that Ganoderma lucidum has abilities to induce neuronal differentiation and prevent NGF-dependent PC12 neuronal cells from apoptosis [35], H.E. mycelium was more effective in neuroprotection than Ganoderma lucidum in this PC12 cells study. It may be due to that fact that a new cell-wall breaking technology was employed and also finer particles were produced. It improves the release of active ingredients from smaller H.E. mycelium particle powders.
By using DPPH radical scavenging assay, widely used method to evaluate the ability of mushroom to scavenge free radicals generated from DPPH reagent [36,37], the present data also showed that H.E. mycelium had more effective in scavenging DPPH radicals when compared to Ganoderma lucidum. The radical inhibition effect of Ganoderma lucidum was 59.3% at concentration of 1000 µg/ml, when compared with H. E. mycelium’s 96.1% and with ascorbic acid’s 100% at the same concentration.
Administration of MPTP was known to decrease the activity of neuron and the density of TH-positive neurons, indicating that the degeneration of the dopaminergic neurons in SNpc [38,39]. In our present study, injection of MPTP into C57BL/6 Narl mice induced significant reduction of dopamine level and TH-positive area in the striatum, suggesting that MPTP initially affects the dopaminergic cell body in the substantia nigra pars compacta (SNpc) and then the striatum where dopaminergic cells exert it’s function of release of dopamine. Tyrosine hydroxylase (TH) is the initial enzyme in the catecholamine synthesis pathway [40] and dopamine biosynthesis in the central nervous system [41]. TH is activated to form more DOPA and then dopamine by decarboxylation which is transferred into the synaptic vesicle by the vesicular monoamine transporter (VMAT). In addition, the loss of TH activity or expression is thought to contribute to dopamine deficiency, which is the most prominent at media level of SNpc (substantia nigra pars compacta) [39]. In our current study, the level of dopamine concentration and immunohistochemistry for TH positive neurons revealed that the loss of dopamine neuron in PD mice was dramatically prevented after treatment of H.E. mycelium.
In consistent with other research, the results of our present study showed that the immuno reactivity of TH was significantly decreased in MPTP-treated mice, suggesting that the majority of dopaminergic neurons were lost in the Parkinson’s disease mouse model [42]. The death of dopaminergic neurons led to decrease dopamine level in the substantia nigra. The expression of TH positive neuron and concentration of dopamine level were increased with the concentration of H.E. mycelium from 0.3 g/kg to 1 g/kg in MPTP-induced mice. This evidence suggested that the neuroprotection and anti-oxidation is involved in the protective effect of H.E. mycelium on dopaminergic neurons.
In conclusion, we have used cell culture and animal models to demonstrate the neuroprotective effect of H.E. myceliumin the PD model. Our results indicate that H.E. mycelium increased the cell viability of the MPP+ treated cells and performed in a very effective antioxidative effect in vitro cell culture model using PC 12 cells. in vivo, H.E. mycelium exerts significant neuroprotective effect by increasing the dopamine level and the activity of tyrosine hydroxylase (TH) in mice striatum. To sum up, our results suggest that H.E. mycelium performs significant protection of dopaminergic neuron under severe conditions and is very effective in the treatment of damaged neuron in the brain to recover in the case of Parkinson’s disease.
Download Provisional PDF Here
Article Type: RESEARCH ARTICLE
Citation: Yang PP, Lin CY, Lin TY, Chiang WC (2020) Hericium erinaceus Mycelium Exerts Neuroprotective Effect in Parkinson’s Disease-in vitro and in vivo Models. J Drug Res Dev 6(1): dx.doi.org/10.16966/2470-1009.150
Copyright: © 2020 Yang PP, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access