
Figure 1: Structure of cellulose butyrate.
Number in parenthesis is the position for substitution by butyl group.

Hiroki Maeoka1 Shu Shimamoto2 Yutaka Suzuki1 Satoshi Koike1 Yasuo Kobayashi1*
1Graduate School of Agriculture, Hokkaido University, Sapporo, Hokkaido, Japan*Corresponding author: Yasuo Kobayashi, Graduate School of Agriculture, Hokkaido University, Kita, Sapporo, Hokkaido 060−8589, Japan, Phone: +81-11-706-2476; E-mail: kyas@agr.hokudai.ac.jp
To evaluate the function of cellulose butyrate (CB), a new synthetic, fibrous, candidate food additive, a series of batch culture study was carried out. Fecal cultures were prepared using fresh feces from human volunteers and swines, and anaerobically incubated to characterize the effect of CB on hindgut fermentation and microbial community, and thereby evaluate the potential use of CB as a food additive in mono-gastric animals. Three types of CB differing in the degree of substitution and in the carbon position of glucose residue where butyl groups are substituted were tested to proceed multiple comparisons for measured parameters. Among three types of CB tested, partially water-soluble CB3 was associated with the highest increase in butyrate production in human and swine fecal cultures. CB3 supplementation shifted fecal microbiota toward greater abundance of bacteria in the Bacteroidetes phylum, in particular the genus Parabacteroides, in which the increase of Parabacteroides distasonis was remarkable even with individual variations. CB3 also lowered putrefactive products, including ammonia, indole and skatole, in the cultures. Dietary use of partially water-soluble CB3 might be expected to have useful effects in humans and other mono-gastric animals, judging from the present results of enhanced butyrate production and lowered putrefactive products with CB3 addition to feces. The results might be partly attributable to a shift in hindgut microbiota, represented by the increased abundance of Bacteroidetes, especially Parabacteroides species. Therefore, CB3 can be considered as a functional additive candidate for the use in mono-gastric animals such as human and pigs.
Fecal culture; Microbial community; Mono-gastric animal; Parabacteroides
The intestinal microbiota of mono-gastric animals plays a pivotal role in digestion and fermentation of dietary and endogenous polysaccharide components that escape digestion by host enzymatic systems, providing short chain fatty acids (SCFAs) represented by acetate, propionate and butyrate through microbial degradation and fermentation [1]. SCFAs provide 10% of the maintenance energy requirements in humans[2], and provide various health benefits [3]. In particular, butyrate confers beneficial effects on human health, serving as an energy source for colonic epithelial tissues [4], promoting the synthesis of epithelial membranes, regulation of cytokines, and in intestinal barrier function [5]. Direct infusion of butyrate to the rectum of mice enhances gene expression of tight junctions and suppresses cell permeability, which prevents pathogenic invasion of the body [6]. Butyrate regulates the expression of interleukin-6 (IL6) and tumor necrosis factor alpha (TNF-α), both of which are known to cause excessive inflammation [7]. Thus, enhancement of butyrate production contributes to functional improvement of the intestinal tissue barrier and suppression of inflammation by related cytokines, possibly leading to better host animal health.
Based on these scientific backgrounds, much attention has been paid to butyrate utilization in the farm animal industry. The high incidence of diarrhea in piglets after weaning is a big concern due to the resultant lowered growth [8]. Although antibiotic administration has been the mainstay of treatment for this issue since many years [9], the use of growth-promoting antibiotics is being reconsidered or even banned in the EU since 2006. Hence, we need new alternative substances to regulate intestinal microbiota and fermentation. One such candidate substance is a butyrate additive. Haung, et al. [10] tested a sodium butyrate additive in weaned piglets (1000mg/kg diet) and reported decreased occurrence of diarrhea (33%) and an increase in body weight gain (10%) with enhancement of intestinal barrier function through improved gene expression of tight junction proteins. They also clarified a shift of intestinal microbiota toward an increase in Ruminococcaceae and Lachnospiraceae, which are considered as beneficial probiotic bacteria that promote energy acquirement from polysaccharides. Thus, the increased abundance of these beneficial bacteria by butyrate additives might be useful for modulation of intestinal microbiota and tissues, promoting host animal health [11,12].
Cellulose butyrate (CB) is a new synthetic fiber with multiple functional uses, among which dietary application to mono-gastric animals including humans, pigs, and poultries can be one of the targets. CB is a cellulose derivative, in which hydroxyl-groups of a glucose residue are partially substituted by a butyl-group. The degree of substitution defines the physical nature of CB, such as its watersolubility, as described in cellulose acetate [13,14]. Use of CB as a food additive in mono-gastric animals is expected to cause an increase in butyrate in the animal’s gastrointestinal (GI) tract, due to release of the butyl-group from CB. Based on the functionality of butyrate described above, increased butyrate production in the GI tract might promote the health of CB-fed animals.
Degradation of CB could be preceded by release of the butyl-group by esterase enzymes, as reported for the release of the acetyl-group from cellulose acetate (CA) [13,14]. Many symbiotic GI bacteria possess various esterase enzymes, as seen by the fact that 950 of 15,862 carbohydrate-active enzymes are esterases [15]. Thus, release of the butyl-group from CB occurs first, followed by degradation of the cellulose main chain. For instance, CA is utilized by specific GI bacteria, including Bacteroides spp., especially Bacteroides uniformis, having esterases, and growth of these bacteria is often seen in conjunction with proliferation of other bacteria involved in hydrolyzation of the cellulose main chain to produce more SCFAs [14]. Although CB is assumed to be utilized by GI bacteria in a manner similar to CA, the rate and extent of SCFA production, in particular butyrate production, needs to be experimentally assessed for deeper understanding of fermentation and the future application of CB.
The present study attempted to evaluate the possible use of CB as a novel food additive candidate for mono-gastric animals, including humans and pigs, through analysis of the fermentability and microbial community in fecal cultures with and without CB addition.
The present study was carried out according to the Guidelines for Animal Experiments, Hokkaido University (2007), and the Act on Welfare and Management of Animals (2005). The study on human stool cultures was approved by the Ethics Committee of the Research Faculty of Agriculture, Hokkaido University (approval number: H301210-No.2).
The basic structure of CB and the structural characteristics of the CBs tested in the present study are shown in figure 1 and table 1, respectively. CB1 and CB2 were insoluble, having 0.9 and 1.4 degrees of substitution of the hydroxyl group by a butyl group (3.0 degrees is the maximum possible substitution, because a glucose residue has 3 hydroxyl groups). The substitution is high at the 6-position (0.5) for CB1, and is high at the 2- and 3-positions (0.6) for CB2. CB3 had the same substitution degree of 0.9 as CB1, but with a lower substitution at the 6-position (0.1), which made this compound partially (approximately 30%) water-soluble. All these CBs were manufactured by Daicel Corporation, Tokyo, Japan. Functions of these CBs were evaluated in 3 in vitro experiments; 1st for initial evaluation using human feces from a single donor, 2nd for evaluation using swine feces, and 3rd for final evaluation using human feces from 4 individual donors.
Figure 1: Structure of cellulose butyrate.
Number in parenthesis is the position for substitution by butyl group.
| Degree of substitution (DS) | |||||
| Material tested | Treatment | Total DS | C2 | C3 | C6 |
| WSCB2015/12 | CB1 | 0.9 | 0.2 | 0.2 | 0.5 |
| NCB-1L-1L | CB2 | 1.4 | 0.6 | 0.6 | 0.2 |
| NCB-1-240-2 | CB3 | 0.9 | 0.4 | 0.4 | 0.1 |
Table 1: Molecular characteristics of cellulose butyrate (CB) used in this
study.
C2, C3, C6; Carbon positions of glucose residue where butyl groups can be substituted.
Experiment 1: Evaluation with human fecal cultures: A fresh stool sample was provided by a 25-year-old healthy male volunteer who had not taken any antibiotics, probiotics and prebiotics but taken rice grain-based diet commonly seen in Japan for the preceding 3 months. The feces was diluted to form a 10% (w/v) fecal slurry with 0.1 M phosphate buffered saline (PBS; 8 g L–1 NaCl, 0.2 g L–1 KCl, 1.15 g L–1 Na2HPO4, 0.2 g L–1 K2HPO4, 8 g L–1 NaCl, 0.2 g L–1 KCl, 1.15 g L–1 Na2HPO4, 0.2 g L–1 KH2PO4). The diluted fecal solution was filtered through two layers of surgical gauze, and 1 ml of the filtrate was added to 9 ml of the intestinal environmental medium (2 g L–1 peptone water, 2 g L–1 yeast extract, 0.1 g L–1 NaCl, 0.04 g L–1 K2HPO4, 0.04 g L–1 KH2PO4, 0.01 g L–1 MgSO4·7H2 O, 0.01 g L–1 CaCl4·6H2 O, 0.5 g L–1 bile salts, 2 mL L–1 Tween 80, 1 mL L–1 of 0.05% hemin solution, 0.01 mL L–1 vitamin K1 , 1 mL L–1 0.1% resazurin sol., 0.5 g L–1 L-cysteine HCl, 2 g L–1 NaHCO3), and used as a fecal culture for the incubation study.
The above fecal culture was dispensed into test tubes in which either of the three different CBs (CB1, CB2 and CB3) had been added (0.1 g/tube). For the control tube, no carbon source was added. The tubes were flushed with nitrogen gas, fitted with a butyl rubber stopper and a plastic screw cap, and then anaerobically incubated at 37℃ for 24 h. Incubation was performed in quadruplicate per treatment. After incubation, the cultures were centrifuged at 13,000 × g at 4℃ for 2 min to obtain a supernatant that was promptly used for pH measurement (F21 pH meter, Horiba, Kyoto, Japan), and then frozen at -30℃ for later analyses of SCFA, ammonia, indole and skatole. The uncentrifuged whole culture was also frozen at -80℃ for analysis of microbes.
Experiment 2: Evaluation with swine fecal cultures: Fresh feces was collected from two adult female pigs (Large White x Landrace) at the Livestock Production Research Facility, Northern Biosphere Field Science Center, Hokkaido University. The pigs had been given a commercial feed (Hybrid 72 M; Hokuren Kumiai, Hokkaido, Japan) formulated with 62% grains, 18% oil seed meal of vegetable origin, 11% chaff and bran, and 9% others (beet pulp, calcium carbonate, molasses, calcium phosphate, salt, glucose, fructo-oligosaccharide syrup, calcium fatty acid) having 14% crude protein, 2.0% crude fat, 10.0% crude fiber, 10% crude ash, and 72.0% total digestible nutrients. The collected feces was equally mixed and 10-fold diluted with 0.1 M PBS to prepare the fecal culture. The tested CBs, incubation procedures and samplings were performed by the same method as described for experiment 1.
Experiment 3: Final evaluation with human fecal cultures from four individuals: This experiment was designed to assess variations in the potency of CB3 by using human stools collected from 4 different individuals. A fresh stool sample was provided by four male volunteers (23 ± 0.25 years old) who had the same dietary background as described in the experiment 1. Preparation of the fecal culture, tested CBs, incubation procedures and samplings were performed in the same manner as above, except that only two CBs (CB2 and CB3) were used and the incubation was performed in duplicate for each individual. For microbial analysis, DNA extracted from the duplicated cultures were equally mixed to represent each individual.
SCFA analysis was performed according to the method described by Yamada et al. [14]. In brief, the thawed culture supernatant was pretreated by mixing with 2℃ propanol (99.7%) at a 4:1 ratio, left overnight at 4℃, and then centrifuged at 10,000 × g at 4℃ for 5 min. Following the addition of an internal standard (crotonic acid) to the supernatant, the mixture was injected into a GC-14B gas chromatograph (Shimadzu, Kyoto, Japan) equipped with an Ulbon HR-20M fused silica capillary column (0.53mm inner diameter × 30m length, 3.0-µm film; Shinwa Chemical Industries, Kyoto, Japan) and a flame-ionization detector. Helium and nitrogen gases were used as the carrier gas and makeup gas, respectively. The concentration of each SCFA in the sample was determined based on its ratio to crotonic acid and the chromatographic pattern of a standard mixture in which the concentration of each SCFA was known.
Ammonia was measured by the indophenol blue method [16], reading the absorbance at 660 nm, while indole and skatole were measured by colorimetric methods at a wave length of 560 nm [17]. Both measurements were carried out using a microplate reader (PerkinElmer Japan Co. Ltd., Kanagawa, Japan).
Extraction of bacterial DNA for the microbial analysis was performed using the repeated bead beating plus column method described by Yu and Morrison [18]. The abundances of total bacteria, Bacteroides-Prevotella-Porphyromonas, Ruminococcaceae, Clostridium coccoides-Eubacterium rectale group, Faecalibacterium prausnitzii, genus Bifidobacterium, Eubacterium hallii, genus Prevotella, Escherichia coli, genus Lactobacillus, Akkermansia muciniphila, and Clostridium perfringens were estimated by quantitative real-time polymerase chain reaction (qPCR) assays with a LightCycler 480 (Roche Applied Science, Penzberg, Germany) and a Kapa SYBR Fast qPCR Kit (Kapa Biosystems, Charlestown, MA, USA). The qPCR was performed as follows: denaturation (95℃ for 3 min), followed by 40 cycles of denaturation (95℃for 10 s), annealing (at the temperature set for the target for 20 s), and extension (72℃ for the time duration set for the target) (see Supplemental table). The relative abundance of a specific bacterial group was shown as a proportion of the total bacterial abundance.
| Target | Primer sequences(5'-3') | Annealing Temperature (℃) |
Extension Time (sec) |
Product Size (bp) | Reference |
| Total bacteria | [F] CCTACGGGAGGCAGCAG [R] ATTACCGCGGCTGCTGG |
60 | 1 | 194 | Muyzer et al., (1993) |
| Bacteroides- Prevotella-Porphyromonas |
[F] GGTGTCGGCTTAAGTGCCAT [R] CGGAYGTAAGGGCCGTGC |
68 | 1 | 140 | Rinttila et al., (2004) |
| Ruminococcaceae | [F] GCACAAGCAGTGGAKT [R] CTTCCTCCGTTTTGTCAA | 50 | 1 | 239 | Matsuki et al., (2004), modified* Matsuki et al., (2004) |
| Clostridium coccoides-Eubacteriumrectalegroup | [F] GCACAAGCAGTGGAGT [R] CTTCCTCCGTTTTGTCAA | 50 | 1 | 239 | Matsuki et al., (2004) |
| Faecalibacteriumprausnitzii | [F] GGAGGAAGAAGGTCTTCGG [R]AATTCCGCCTACCTCTGCACT | 60 | 1 | 248 | Ramirez et al., (2009) |
| Genus Bifidobacterium | [F] GGGTGGTAATGCCGGATG [R] CCACCGTTACACCGGGAA | 62 | 2 | 523 | Kok et al., (1996) |
| Eubacteriumhallii | [F] GCGTAGGTGGCAGTGCAA [R] GCACCGRAGCCTATACGG | 60 | 1 | 278 | Ramirez et al., (2009) |
| Genus Prevotella | [F] CACRGTAAACGATGGATGCC [R] GGTCGGGTTGCAGACC | 55 | 2 | 534 | Matsuki et al., (2002) |
| Genus Lactobacillus | [F] AGCAGTAGGGAATCTTCCA [R] CACCGCTACACATGGAG | 58 | 1 | 371 | Walter et al., (2001) Heiling et al., (2002) |
| Akkermansiamuciniphila | [F]CAGCACGTGAAGGTGGGGAC [R] CCTTGCGGTTGGCTTCAGAT | 60 | 1 | 327 | Png et al., (2010) |
| Clostridium perfringens | [F]AAAGATGGCATCATCATTCAAC [R]TACCGTCATTATCTTCCCCAAA | 55 | 1 | 279 | Wang et al., (1994) |
Supplementary table: Primers used in this study for real-time PCR.
*:[F] GCACAAGCAGTGGAGT in Matsuki et al.(2004) was modified to [F]GCACAAGCAGTGGAKT.
To analyze the microbial community more comprehensively, MiSeq analysis (Illumina, Inc., San Diego, CA, USA) was performed using the samples of experiment 3. The V3 to V4 regions of the 16S rRNA gene were amplified using the following primers: S-D-Bact-0341-b-S-17 (5′‐CCTACGGGNGGCWGCAG-3′) and S-D-Bact0785-a-A-21 (5′‐ GACTACHVGGGTATCT AATCC-3′) [19]. The resulting libraries were sequenced using the MiSeq Reagent Kit V3 [20] according to Illumina’s 2 × 300-bp paired-end sequencing protocol. Data quality control and analyses were performed using the QIIME2 2018.11 program [21]. The DADA2 plug-in [22] was used to control data quality, following which the feature table was generated. Taxonomy was assigned using the Greengenes database (version 13.8) [23] at a 90% similarity threshold. Differences in biodiversity between control and treatment samples were compared by alpha diversity metrics, including the observed number of operational taxonomic units (OTU), Good’s coverage, Chao1, Shannon index, and Simpson index.
The de-butylation rate was assumed by dividing the net production of butyrate (butyrate production in CB-butyrate production in control) by the theoretically maximal amount of butyrate that can be liberated from CB by de-butylation with esterases. Bacterial group was expressed as the relative % of total bacteria dividing the value of each group by that of total bacteria both in qPCR and MiSeq data. Groups at lower than quantifiable range of qPCR were expressed as “not detected”
Data in the tables are expressed as means with pooled standard errors of the mean (SEMs). Data were analyzed by one-way ANOVA, and significant differences were judged by multiple comparisons (Tukey-Kramer method) with GraphPad Prism 5 (GraphPad Software, San Diego, USA). Statistical significance was inferred at P<0.05.
The effects of CB on pH, SCFA and putrefactive products in human fecal culture are shown in table 2. CB3 increased total SCFA, acetate, propionate and n-butyrate concentrations, leading to a lower pH. CB2 also increased n-butyrate concentration, but not the other SCFAs. In terms of molar proportion, n-butyrate was elevated by CB2 and, in particular, CB3, exceeding propionate to be the second major SCFA in the culture. The calculated de-butylation rates for CB2 and CB3 were 1.3 and 13.1%, respectively, while this value was quite low (0.3%) for CB1. Ammonia level was lowered by CB3 in comparison with control and CB2. Indole and skatole concentrations decreased with CB2 and CB3 in comparison with control and CB1.
| Parameter | Control | CB1 | CB2 | CB3 | Pooled SEM |
| pH | 6.44b | 6.50a | 6.47ab | 5.54c | 0.006 |
| Total short chain fatty acid, mmol L-1 | 20.1b | 21.2b | 22.0b | 43.7a | 0.32 |
| Acetate, mmol L-1 | 13.5b | 14.2b | 14.5b | 29.4a | 0.21 |
| Propionate, mmol L-1 | 2.8b | 2.9b | 3.0b | 5.7a | 0.04 |
| iso-Butyrate, mmol L-1 | 0.7ab | 0.8a | 0.7a | 0.6b | 0.02 |
| n-Butyrate, mmol L-1 | 1.9c | 1.9c | 2.5b | 7.0a | 0.06 |
| iso-Valarate, mmol L-1 | 1.3a | 1.5a | 1.4a | 1.1b | 0.03 |
| Valarate, mmol L-1 | ND | ND | ND | ND | ND |
| Acetate, molar% | 67.1 | 66.8 | 65.6 | 67.2 | 0.23 |
| Propionate, molar% | 13.9a | 13.6ab | 13.6ab | 13.0b | 0.10 |
| iso-Butyrate, molar% | 3.4ab | 3.8a | 3.3b | 1.3c | 0.07 |
| n-Butyrate, molar% | 9.2c | 9.0d | 11.3b | 16.0a | 0.10 |
| iso-Valarate, molar% | 6.4ab | 6.8a | 6.2b | 2.5c | 0.09 |
| Valarate, molar% | ND | ND | ND | ND | ND |
| NH4 -N, mg L-1 | 38.3a | 36.1ab | 37.8a | 35.8b | 0.16 |
| Indole, µg ml-1 | 0.9a | 0.8a | 0.3b | 0.3b | 0.02 |
| Skatole, µg ml-1 | 2.9a | 3.3a | 1.0b | 1.8b | 0.16 |
Table 2: Effect of cellulose butyrate (CB) supplementation on fermentation
parameters in in vitro human fecal cultures.
Values with different letters are significantly
different (P<0.05).
ND, not detected.
Effects of CB on pH, SCFA and putrefactive products in swine fecal cultures are shown in table 3. The changes in all the parameters determined were almost similar to those observed in human fecal cultures. The pH was lower in CB3 than in control and other CBs, along with higher total SCFA concentration in comparison with control and other CBs. In terms of molar proportion, n-butyrate became higher with CB3 than with the other CBs. In comparison with controls, ammonia level was lowered by CB3, while remaining unchanged by CB1 and CB2.
| Parameter | Control | CB1 | CB2 | CB3 | Pooled SEM |
| pH | 6.55b | 6.67a | 6.58b | 5.95c | 0.013 |
| Total short chain fatty acid, mmol L-1 | 19.7b | 17.3b | 18.8b | 49.8a | 0.94 |
| Acetate, mmol L-1 | 10.9b | 9.9b | 10.7b | 29.3a | 0.60 |
| Propionate, mmol L-1 | 2.7b | 2.4b | 2.7b | 7.5a | 0.13 |
| iso-Butyrate, mmol L-1 | 0.7 | 0.6 | 0.5 | 0.7 | 0.03 |
| n-Butyrate, mmol L-1 | 2.9b | 2.4b | 2.8b | 10.0a | 0.18 |
| iso-Valarate, mmol L-1 | 1.4 | 1.2 | 1.0 | 1.2 | 0.05 |
| Valarate, mmol L-1 | 1.1 | 0.8 | 1.2 | 1.1 | 0.08 |
| Acetate, molar% | 55 | 57.1 | 56.9 | 58.8 | 0.55 |
| Propionate, molar% | 13.9 | 13.8 | 14.2 | 15.1 | 0.23 |
| iso-Butyrate, molar% | 3.7a | 3.5a | 2.5ab | 1.4b | 0.17 |
| n-Butyrate, molar% | 14.9b | 14.0b | 14.8b | 20.0a | 0.23 |
| iso-Valarate, molar% | 7.0a | 7.0a | 5.4a | 2.5b | 0.24 |
| Valarate, molar% | 5.5ab | 4.6b | 6.2a | 2.2c | 0.21 |
| NH4 -N, mg L-1 | 29.0a | 27.7ab | 29.5a | 24.4b | 0.56 |
| Indole, µg ml-1 | 0.9ab | 1.1a | 0.5b | 0.5b | 0.07 |
| Skatole, µg ml-1 | 1.4ab | 2.1a | 1.1ab | 0.7b | 0.17 |
Table 3: Effect of cellulose butyrate (CB) supplementation on fermentation
parameters in in vitro porcine fecal culture.
Values with different letters are significantly different (P<0.05).
Relative abundances of representative bacterial species/groups in human and swine fecal cultures as influenced by CBs are shown in figure 2. In human fecal cultures, CB3 increased the relative abundances of the Bacteroides-Prevotella-Porphyromonas group and genus Prevotella, compared with controls, while decreasing those of genus Bifidobacterium, the C. coccoides-Eubacterium rectale group and F. prausnitzii in comparison with controls. The other two CBs did not alter the relative abundance of bacterial groups, except that CB2 decreased the abundance of genus Bifidobacterium and F. prausnitzii. In porcine fecal cultures, almost similar changes were observed with CB3, which increased the relative abundance of the Bacteroides-Prevotella-Porphyromonas group, while decreasing those of the C. coccoides-Eubacterium rectale group and F. prausnitzii. CB3 also increased the abundance of genus Lactobacillus, and decreased that of Ruminococcaceae. Other CBs did not change the microbiota specifically, except for an increase in Bacteroides-PrevotellaPorphyromonas group by CB2 and some other minor changes.
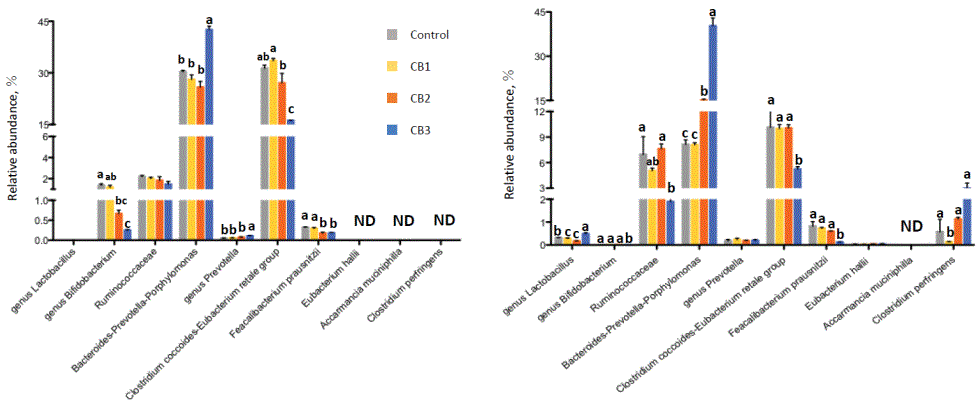
Figure 2: Effect of cellulose butyrate (CB) supplementation on bacterial groups in human (upper) and porcine (lower) fecal cultures determined by real-time PCR assays.
No carbohydrate was supplemented for control, while either of 3 different CBs was supplemented for CB1, CB2, and CB3.
Values with different letters are significantly different (P<0.05).
ND, not detected.
The effects of CB2 and CB3 on pH, SCFA and putrefactive products in fecal cultures prepared from four individuals are shown in table 4. The pH was decreased, while all three major SCFAs (acetate, propionate and n-butyrate) were enhanced by CB3 in comparison with control, even though molar proportions of SCFA did not differ. CB2 did not change SCFA levels and proportions. The calculated de-butylation rate was 8.5% for CB3, while that of CB2 was 0.6%. Ammonia concentration did not change with CBs, while indole and skatole levels were lowered by both CB2 and CB3 in comparison with controls.
| Parameter | Control | CB2 | CB3 | Pooled SEM |
| pH | 6.54b | 6.68a | 5.86c | 0.026 |
| Total short chain fatty acid, mmol L-1 | 18.2b | 20.3b | 36.1a | 0.64 |
| Acetate, mmol L-1 | 12.5b | 13.8b | 25.2a | 0.33 |
| Propionate, mmol L-1 | 2.6b | 2.7b | 4.8a | 0.26 |
| iso-Butyrate, mmol L-1 | 0.3 | 0.5 | 0.3 | 0.08 |
| n-Butyrate, mmol L-1 | 2.0b | 2.3b | 5.4a | 0.35 |
| iso-Valarate, mmol L-1 | 0.6 | 0.8 | 0.4 | 0.16 |
| Valarate, mmol L-1 | 0.2 | 0.2 | 0.1 | 0.13 |
| Acetate, molar% | 68.7 | 67.9 | 69.9 | 1.06 |
| Propionate, molar% | 14.2 | 13.5 | 13.3 | 0.91 |
| iso-Butyrate, molar% | 1.8 | 2.3 | 0.8 | 0.33 |
| n-Butyrate, molar% | 10.9 | 11.3 | 14.8 | 1.01 |
| iso-Valarate, molar% | 3.6 | 3.9 | 1.1 | 0.74 |
| Valarate, molar% | 0.9 | 1.1 | 0.2 | 0.62 |
| NH4 -N, mg L-1 | 29.0ab | 35.9a | 22.4b | 2.46 |
| Indole, µg ml-1 | 1.4a | 0.9b | 1.0b | 0.03 |
| Skatole, µg ml-1 | 5.0a | 2.8b | 3.4b | 0.40 |
Table 4: Effect of cellulose butyrate supplementation on fermentation
parameters in human fecal cultures prepared from 4 individuals.
Values with different letters are significantly different (P<0.05).
Figure 3 shows the relative abundance of bacterial species/groups as influenced by CBs. Similar changes to those by CB3 in experiment 1 were observed (increased Bacteroides-Prevotella-Porphyromonas group and decreased C. coccoides-E. rectale group). On the other hand, CB2 did not show any particular changes in comparison with controls.
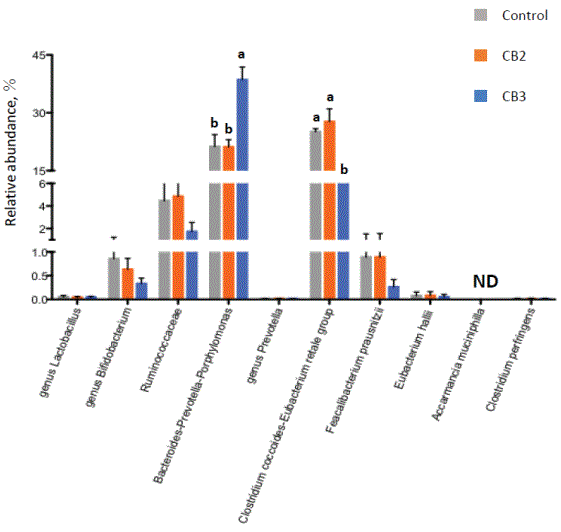
Figure 3: Effect of cellulose butyrate (CB) supplementation on bacterial abundance in human fecal cultures prepared from 4 individuals.
No carbohydrate was supplemented for control, while either of 2 different CBs was supplemented for CB2 and CB3.
Values with different letters are significantly different (P<0.05).
ND, not detected.
Diversity indices of bacteria in fecal cultures by MiSeq analysis are shown in table 5. The number of sequences read were 22,953, 22,994 and 22,731 for control, CB2 and CB3, respectively. No significant differences between treatments were observed for all diversity indices, including the operational taxonomic unit (OTU) number, Good’s coverage, Chao 1, Shannon and Simpson indices.
| Control | CB2 | CB3 | Pooled SEM | |
| Operational taxonomic units | 121 | 126 | 101 | 17.14 |
| Good’s coverage, % | 100 | 100 | 100 | - |
| Chao1 | 120.5 | 126.0 | 100.5 | 17.14 |
| Shannon | 5.12 | 5.21 | 4.61 | 0.16 |
| Simpson | 0.94 | 0.94 | 0.92 | 0.01 |
Table 5: Effect of cellulose butyrate (CB) supplementation on bacterial diversity in human fecal cultures from 4 individuals determined by MiSeq analysis.
Bacteria detected at a 1% or higher level by MiSeq analysis are shown at the genus level in table 6. The relative abundance of Parabacteroides was higher with CB3 than in controls, while unclassified Enterobacteriaceae were lower in the CB3 than control and CB2 samples. The genus Bacteroides increased numerically, although the increase had no statistical significance. When attempting classification at the species level, Bacteroides uniformis and Parabacteroides distasonis showed an increase (B. uniformis, 4.7 vs 2.2 ver 18.5%; and P. distasonis, 2.5 vs 6.4 vs 19.7%, for control vs CB2 vs CB3, respectively). Although these differences were not significant due to individual variations, there were two types of bacterial shifts depending on the individual; three of four individuals tested showed an increase in Parabacteroides distasonis with CB3, while one showed an increase of Bacteroides uniformis (Figure 4).
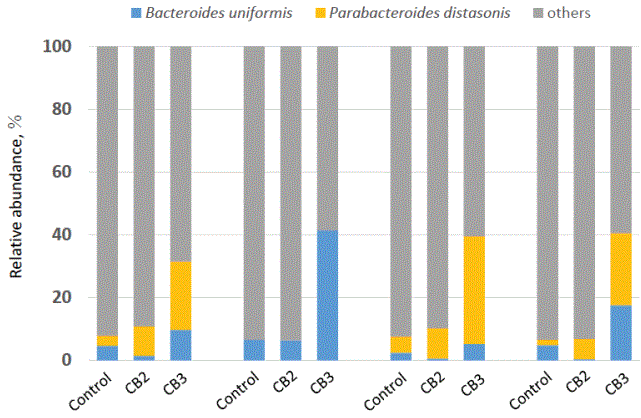
Figure 4: Effect of cellulose butyrate (CB) supplementation on Bacteroides and Parabacteroides species in human fecal cultures from 4 individuals.
No carbohydrate was supplemented for control, while either 2 different CBs was supplemented for CB2 and CB3.
Values with different letters are significantly different (P<0.05).
| Relative abundance, % | Control | CB2 | CB3 | pooled SEM |
| Bacteroides | 26.6 | 21.0 | 33.0 | 3.79 |
| Unclassified Enterobacteriaceae | 15.7a | 16.4a | 5.3b | 1.84 |
| Unclassified Lachnospiraceae ① | 7.2 | 6.6 | 3.7 | 1.23 |
| Clostridium(Lachnospiraceae) | 6.9 | 7.3 | 3.6 | 1.27 |
| Parabacteroides | 6.6b | 11.4ab | 22.4a | 2.63 |
| Oscillospira | 5.9 | 5.7 | 2.2 | 2.31 |
| Veillonella | 5.2 | 4.7 | 5.3 | 3.70 |
| Phascolarctobacterium | 4.7 | 4.0 | 1.9 | 1.88 |
| Blautia | 1.9 | 1.9 | 1.5 | 0.34 |
| [Ruminococcus] (Lachnosperaceae) | 1.8 | 2.0 | 1.4 | 0.33 |
| Unclassified Lachnospiraceae ② | 1.7 | 1.6 | 0.4 | 0.31 |
| Fusobacterium | 1.7 | 2.2 | 0.2 | 0.71 |
| Dorea | 1.4 | 1.7 | 0.3 | 0.40 |
| Sutterella | 1.2 | 1.4 | 0.4 | 0.37 |
| Bifidobacterium | 1.2 | 1.0 | 0.8 | 0.29 |
| Faecalibacterium | 1.1 | 0.6 | 0.3 | 0.39 |
| Dialister | 1.0 | 1.1 | 1.2 | 0.75 |
| Ruminococcus(Ruminococcaceae) | 0.8 | 1.0 | 0.8 | 0.37 |
| Collinsella | 0.4 | 0.3 | 2.0 | 0.81 |
| Clostridium(Erysipelotrichaceae) | 0.2 | 0.2 | 6.8 | 1.15 |
Table 6: Effect of cellulose butyrate (CB) supplementation on fecal
bacterial community at genus level determined by MiSeq analysis.
Values with different letters are significantly different (P<0.05).
Genera exceeding 1% in relative abundance are shown in the alignment with values in control.
CB3 addition increased SCFA production, in particular n-butyrate production, in human and swine fecal cultures (Tables 2 & 3), suggesting that CB3 is hydrolysable and fermentable in their hindgut environments. The first step in CB fermentation is thought to be release of the butyl group from CB by esterases, as seen with CA[14,24]. When the liberation rate of the butyl group was calculated by dividing actually measured butyrate by maximally releasable butyrate from CB3 in the culture, the rate ranged from 8.5 (experiment 2) to 13.1% (experiment 1). Insoluble CBs showed much lower liberation rates, suggesting the importance of water-solubility for access of the ester bonds of CB to esterase enzymes, as has been shown for CA [14]. The water solubility of CB can be defined by the physico-chemical structure of CB, in terms of what degree and what position of the hydroxyl group in the glucose residue is substituted by a butyl group. Although CB3 is not completely water-soluble, it might be much more fermentable in the hindgut of humans, pigs and other mono-gastric animals, compared with insoluble CBs. Keys to increasing the water-solubility of CB could be the degree of substitution, and also the positions for substitution. A high degree of substitution by the acetyl group [14,25], especially at the 6-position [26], hampers access of esterase enzymes to CA through steric hindrance. In fact, acetyl esterase shows a lower activity for the acetyl group at the 6-position of CA, compared with those at the 2- and 3-positions [24].
In fact, CB3, which has a low degree of substitution at the 6-position (0.1) (Table 1), is thought to allow easy access of esterase enzymes to their target sites. This could lead to release of the butyl group followed by greater fermentation of the main cellulose chain, producing not only butyrate, but also acetate and propionate (Tables 2-4). In contrast, none of the major SCFAs was increased by CB1, which has the same degree of substitution but with higher substitution at the 6-position, which causes structural hindrance for enzyme access, minimizing liberation of the butyl group, degradation and fermentation of the main cellulose chain.
Increased SCFAs with CB3 supplementation, especially butyrate, can contribute to the energy supply of host animals and play many other roles as regulators of epithelial generation, gut barrier function and cytokine production, all of which lead to maintenance of the animal’s health [5]. SCFAs produced in the hindgut of humans and pigs account for 10% [2] and 30% [27] of their maintenance energy supply, respectively. Besides, acetate and propionate, both of which are increased by CB3, are involved in health maintenance; acetate works in promotion of the immune system, symptomatic relief of enteritis, and prevention of diabetes [28-31], while propionate serves as a glucogenic substance [32]. All these beneficial effects of SCFAs can be expected if CB3 or other more potent CBs are used as food additives (or a postbiotics) in the future.
Hindgut putrefactive fermentation products, represented by ammonia, indole and skatole, were decreased by CBs, in particular CB3, in human and porcine fecal cultures (Tables 2-4). These are harmful products, causing apoptosis-inducing intestinal inflammation [33]. Reduction of these harmful products by CBs could contribute to the maintenance or promotion of gut health in mono-gastric animals. In addition, possible minimization of these odorous products by CB feeding can be a good strategy for the pig industry, and also for mono-gastric companion animals kept indoors for a relatively long time. Reduction of these putrefactive products with CB3 could partly be attributed to the shift in microbiota, such as the decrease of Ruminococcaceaae (Figure 2) and Clostridium (Table 6); some species of these groups are involved in tryptophan metabolism, leading to the formation of indole and skatole [34]. The decrease in unclassified Enterobacteriaceae (Table 6), including pathogenic bacteria, with CB3 supplementation might also contribute to improving gut environment [35].
The most significant change in fecal microbiota by CBs was the increase in Parabacteroides (6.6, 11.4 and 22.4% for controls, CB2 and CB3, respectively, Table 6). Although this is not the same as observed for CA (which is associated with an increase in Bacteroides) [14], Bacteroides also tended to increase with CB3. However, the increase was not statistically significant due to individual variations. Of these two genera, Parabacteroides distasonis and Bacteroides uniformis were found to clearly increase with CB3 supplementation, depending on the individual (Figure 4). This indicates that the response of fecal microbiota to CB3 can be separated into two types, one for increased P. distasonis, and the other for increased B. uniformis. This separation might be due to the difference in indigenous microbial community structure between the individuals from whom fecal samples were obtained.
P. distasonis has been reclassified from Bacteroides to Parabacteroides, which has CE4 family enzymes, comprising more than 7,600 enzymes, including polysaccharide deacetylase, such as acetyl-xylan esterase [36]. Meanwhile, B. uniformis possesses polysaccharide utilization loci, in which CE7 family enzymes, including acetyl esterase and acetyl-xylan esterase are encoded [37]. Therefore, it is reasonable that these two species are activated to degrade CBs, in which esterases play a pivotal role. In fact, since a high correlation (P<0.01) was detected between butyrate level and the sum of relative abundance of P. distasonis and B. uniformis, these two species might be key groups to produce butyrate by CB.
Furthermore, the importance of P. distasonis in suppression of toxigenic intestinal inflammation in mice has been pointed out by Koh, et al. [38], who found that the membrane fragment of P. distasonis can depress the receptor activity for inflammatory cytokines induced by feeding a high fat diet, and lead to apoptosis of cancer cells. Thus, this bacterial species might be one of the beneficial bacteria in terms of maintenance of health in humans and other long-lived mono-gastric animals.
In conclusion, among the CBs evaluated in the present in vitro fecal cultures, CB3 was found most degradable and fermentable for supplying SCFA, in particular n-butyrate. The extent of fermentation of CB depends on its physico-chemical structure; low substitution of the butyl group at the 6-position of the glucose residue is preferable for higher fermentability. CB3 lowers putrefactive products, which is another beneficial characteristic for maintaining and promoting hindgut health in humans and other mono-gastric animals. All of these functions, however, require more detailed evaluation using feeding experiments. Moreover, CB still has room for improvement, because it is not perfectly soluble and hence, its de-butylation rate is limited. More potent CBs should be designed and evaluated to increase their applicability in future.
The authors are grateful to the staff of Field Science Center, Hokkaido University for collection of fecal samples from finishing pigs.
We certify that there is no conflict of interest related to the present study.
Download Provisional PDF Here
Article Type: RESEARCH ARTICLE
Citation: Maeoka H, Shimamoto S, Suzuki Y, Koike S, Kobayashi Y (2023) Cellulose Butyrate as a Potent Fiber Supplement for Mono-Gastric Animals to Modulate Hindgut Microbiota and Fermentation: In Vitro Evaluations. J Anim Sci Res 7(1): dx.doi.org/10.16966/2576-6457.164
Copyright: © 2023 Maeoka H, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access