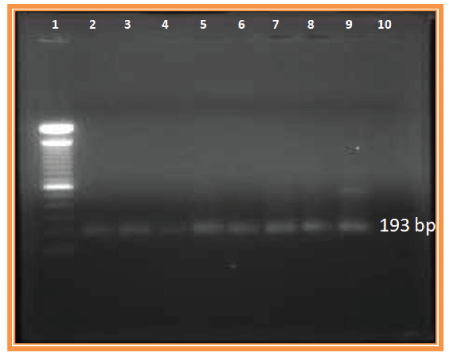
Figure 1: Agarose Gel electrophoresis of PCR products of V3 region Lane 1: 100 bp DNA Ladder; Lane 2 to 9: PCR product (193 bp).

Kiruthiga R1 Thanislass J1* Antony PX2 Lydia DC3 Uma Maheswari D4 Venkatesa Perumal S1
1Department of Veterinary Biochemistry, Rajiv Gandhi Institute of Veterinary Education and Research, Kurumbapet, Puducherry, India*Corresponding author: Thanislass J, Department of Veterinary Biochemistry, Rajiv Gandhi Institute of Veterinary Education and Research, Kurumbapet, Puducherry, India, Tel: +91-413-2273001; Fax: +91- 413- 2272005; E-mail: jthanislass@gmail.com
Molecular ecology techniques based on small subunit (SSU) rRNA sequences can potentially provide a complete description of the rumen microbes. The present study describes the PCRSSCP analysis of V3 region of 16S rRNA as a potential target to characterize the rumen microbiome using metagenomic DNA. All the hundred DNA samples isolated from the goat rumen liquor were subjected to amplification of 16S rRNA gene using a Bacterial universal primer. The V3 region of 16S rRNA was amplified using first round PCR product as template. Sixty-three samples showed positive for amplification of V3 region. Those PCR products were screened for SSCP and found eleven different SSCP patterns. Five representative samples which had produced five different SSCP pattern were sequenced and subjected to sequence analysis. The sequence analysis of V3 region and phylogenetic tree indicated that the DNA sequence of V3 region was found to cluster differently and found to be in association with the SSCP profile of V3 region. Thus the V3 region of 16S rRNA truly reflected the rumen bacterial community. Therefore, it is recommended that the PCR-SSCP of V3 region as an alternative to study the bacterial diversity and its identification in the rumen in place of costintensive metagenomic approaches.
Rumen Microbiome; PCR-SSCP; 16s rRNA; V3 Region; DNA finger Prints
Rumen microbes are essential for ruminant production. A principle function of rumen microbiome is the conversion of plant materials into food products of milk and meat [1]. Therefore rumen is being manipulated to improve the animal production. To manipulate the rumen microbes complete picture of rumen microbiome is important, but it is lacking. Numerous efforts are on to study the genetic diversity of rumen microorganisms and rumen ecosystem. Until recently, knowledge of rumen microbiology was primarily obtained using classical culture-based techniques, such as isolation, enumeration and nutritional characterization which account for 10 to 20% of the rumen microbial population. The methods employed are laborious and time consuming. Efforts are on to identify the culture-independent studies to characterize the rumen microbiome. One such culture-independent study is metagenomics where the microbiome can be accessed by analyzing the collective genomes of the microbial community. New DNA based technologies using metagenomic DNA are now being employed to examine the microbial diversity primarily through the use of small subunit rDNA analysis and to understand the function of the complex microbial ecosystem. The 16S rRNA is universal in bacteria, the conserved region is common for all bacteria and the variable region is unique to particular bacterial genera or species.
Single strand conformation polymorphism (SSCP) is an electrophoretic technique. Under nondenaturing conditions, single strand DNAs will fold into secondary structures (conformations) according to their nucleotide sequences and their physicochemical environment. In nondenaturing conditions, the folded structure of single-stranded DNA (ssDNA) is affected by intra-molecular interactions. The electrophoretic mobility of the folded ssDNA is, therefore, affected by these sequence-dependent properties and the molecular mass of the molecule, allowing separation of PCR products of the same size but different sequence due to the differing mobility of their folded structure [2]. Thus, this technique has been successfully applied to identify sequence variations in a number of genes from different organisms.
Therefore, the PCR amplification of a variable region of 16S rRNA of metagenomic DNA isolated from rumen combined with single strand conformation polymorphism (SSCP) had been applied for the analysis of rumen microbial communities.
Approximately 50ml of Rumen liquor was collected in clean collection bottles from 100 goats slaughtered at Puducherry Municipality Slaughter House. It was transferred via muslin cloth to remove the particulate matter. The filtrate was used for the DNA extraction. Total microbial DNA was extracted by cell lysis method with modification [3]. The quality of DNA was checked by 1% agarose gel electrophoresis and DNA yield was evaluated using spectrophotometer (A260/A280).
The primers which were used for the amplification of bacterial 16S rRNA gene and V3 region of 16S rRNA are given in table 1. Each PCR was carried out in a total volume of 20 µl reaction mixture consisted of 10 pmol of each primer, 2 µg of template DNA and 10 µl of 2x PCR master mix. The PCR amplification of 16S rRNA was performed by thermal cycler (Mastercycler EP, Eppendorf) using the program: initial denaturation of 10 min at 94℃, followed by 30 cycles of denaturation at 94℃ for 60 s, annealing at 50℃ for 60 s and elongation at 72℃ for 60 sec with a final extension at 72℃ for 5 min. The first round PCR product was used as template DNA for second round PCR targeting V3 region of 16S rRNA. Amplification of V3 region was performed using touchdown method which consisted of an initial denaturation at 94℃ for 4 min, 15 cycles of denaturation at 94℃ for 1 min, annealing at 65℃ for 1 min and extension at 72℃ for 1 min in which the annealing temperature was reduced by 0.5℃/cycle from the proceeding cycle and the 15 cycles of 94℃ for 1 min, 55℃ for 1 min and 72℃ for 1 min followed by final elongation at 72℃ for 4 min. The PCR product size (193bp) was checked on 1% agarose gel.
| Primer | Primer sequence (5’à3’) |
Target | Size of the target | Reference |
| 27F | AGAGTTTGATCCTGGCTCAG | Bacterial universal primer 16S rRNA | 1400 bp | Lane DJ et al. [4] |
| 1492R | GGTTACCTTGTTACGAACTT | |||
| V3 F | CCTACGGGAGGCAGCAG | V3 | 193 bp | Muyzer et al. [5] |
| V3 R | ATTACCGCGGCTGCTGG |
Table 1: Details of Primers.
The denaturing loading buffer (10 µl) contains 95% formamide, 10 mM NaOH and 0.025% bromophenol blue was added to 10 µl PCR products. The mixture was heated at 94℃ for 10 minutes and snaps chilled on ice for another 10 minutes. SSCP analysis was carried out as follows: Polyacrylamide gel of 12% concentration, 16cm × 18cm × 1mm size was prepared. The gel was subjected for pre-run at 250 V for 30 minutes at 25℃ using 0.5 × TBE buffer. Denatured sample (20 µl) was loaded into the wells. Electrophoresis was performed at 250 V for 3 hours at 25℃ using 0.5x TBE buffer. According to the procedure of Bassam BJ, et al. [6], the gel was silver stained and the bands were visualized under white light background and documented using gel documentation (Bio-Rad) system.
Five samples which are associated with distinct SSCP pattern were subjected for re-amplification of V3 region and the PCR products were custom sequenced (Eurofins Genomics Pvt Ltd, Bangalore).
The similarity searches for sequences were carried by BLAST. Lineage and Taxonomy report for each sequence is generated from BLAST analysis for the identification of bacteria. The phylogenetic tree was constructed using MOLE-BLAST programme.
Understanding the functions and composition of rumen microbial communities is necessary to improve the animal production [7] and to reduce the methane emission [8]. The conventional culture-based methods for enumerating rumen microorganisms are being replaced by molecular techniques which are being used for the characterization of complex microbial communities and their interactions in the rumen.
Molecular techniques based on amplification of ribosomal genes have allowed quantitative and qualitative studies on microbial populations in the rumen. In prokaryotes, the fulllength 16S rRNA (about 1500 bp) can be used for Taxonomy identification [9] and bacterial diversity [10]. The 16S rRNA sequences consist of nine hypervariable regions (V1-V9) that are separated by nine highly conserved regions [11]. The nine well-defined hypervariable region (V1 to V9) of the 16S rRNA gene contains various taxonomic information [12-14]. Primer pairs targeting on V1 to V3, V3to V5 and V6 to V9 generally result in overall similar and yet accurate classification with minor bias [13]. Most of the studies targeted the V3 region for identification of bacteria [15]. So in this study also, the V3 region of 16S rRNA was selected for the investigation of SSCP pattern and its association with bacterial diversity. PCR-SSCP analysis has allowed the detection of a greater range of microorganisms in environments including the soil [16], drinking water supply system [17], and environmental dust [18].
Out of 100 samples, a total of 63 samples showed amplification of V3 region in the second round PCR. The expected product size 193 bp was checked by 1% agarose gel electrophoresis shown in figure 1. PCR amplified products of DNA samples (63) obtained using a primer set for V3 region were subjected to SSCP analysis. Silver-stained polyacrylamide gel revealed single strand conformational polymorphism pattern. In the sixty-three (63) samples screened, eleven (11) different SSCP patterns were detected, of which, five patterns are frequently seen (indicated by the arrow mark) (Figure 2).
Figure 1: Agarose Gel electrophoresis of PCR products of V3 region Lane 1: 100 bp DNA Ladder; Lane 2 to 9: PCR product (193 bp).
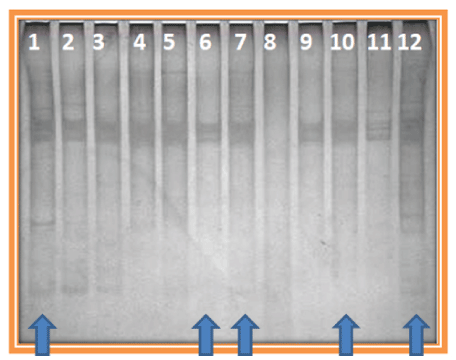
Figure 2: SSCP Pattern of PCR amplified V3 Region of 16S rRNA
The samples (5) associated with the most frequently observed SSCP pattern were selected and amplified the V3 region. The amplified products were sequenced and subjected to sequence analysis. This sequence analysis resulted in the identification of sequence similarity with the bacterial 16S rRNA to the extent of 94%.
Culture-independent 16S rRNA gene sequencing has been widely applied to examine microbial diversity in the chicken gut [19], human gut [20] and identification of bacterial pathogen in clinical samples [21].
The sequence diversity among different bacterial species has meant that 16S ribosomal RNA gene sequences have been widely used for phylogenetic studies and taxonomic classification [22,10]. Due to cost and time management, the partial rrs gene sequences are typically determined in most studies that examined the diversity and species richness of various microbiome. The sequence analysis of V3 [23,24] region of 16S rRNA gene was used for analysis of rumen bacterial diversity. Therefore Sanger sequencing of V3 region was used to characterize the bacterial population in goat rumen.
The phylogenetic tree constructed (MOLE-BLAST) based on the sequence similarity of V3 region had revealed that the clustering of sequences belong to five different samples differently (Figure 3). Differences in a PCR-SSCP profile obtained were found to be associated with a difference in sequences which supports the significance of V3 region in the study of bacterial diversity. The hypervariable V3 region of the 16S ribosomal RNA gene was targeted because it contains the maximum nucleotide heterogeneity that has been selected for the analysis of microbiota [25-27].
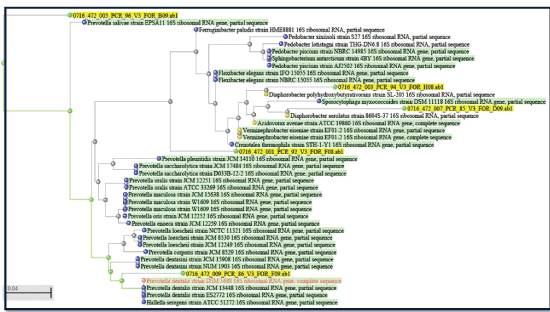
Figure 3: Phylogenetic tree constructed based on sequences of V3 region of 16s rRNA. All the five sequences tested clustered differently.
Based on BLAST analysis, Lineage and Taxonomy reports were generated. The Lineage and Taxonomy report of V3 region have identified the genera Prevotella, Sphingobacterium, Pedobacter, Flavobacterium and uncultured Bacteroidetes belong to Phylum Bacteroidetes. But the Sequence analysis of V4-V5 region identified phylum Proteobacteria contains genera Lysobacter, Aquimonas, Pseudovibrio, uncultured Candidatus, uncultured Burkholderials and the Phylum Bacteroidetes contains the genera Prevotella, Sphingobacterium, Flavobacterium, Croceibacter. The organisms identified based on V3 region are consistent with earlier reports. Bacteroidetes was second most predominant phylum in goat rumen [10]. The genus Prevotella is predominant in the rumen in general [28- 30] reported that the phylum Bacteroidetes was significantly higher in the older animals and composed almost exclusively of the genus Prevotella. The phylum Bacteroidetes in the 110-day old group was significantly higher than in the 90-day old group of goats [1] table 2.
| S.No | Sample No | Phylum | Organisms showing sequence similarity | Function |
| 1 | 85 | Bacteroidetes | Flavobacterium, Sphingobacterium, Bacteroidetes, Pedobacter, Hymenobacter | Cellulose and hemicellulose degradation |
| 2 | 86 | Bacteroidetes | Prevotella, Bacteroidetes | Fiber degradation, Digest the macromolecules |
| 3 | 92 | Bacteroidetes | Prevotella, Bacteroidetes | Fiber degradation Digest the macromolecules |
| 4 | 94 | Bacteroidetes | Prevotella, Sphingobacterium | Cellulose degradation. Fiber degradation |
| 5 | 96 | Bacteroidetes | Prevotella, Bacteroidetes | Fiber degradation. Digest the macromolecules |
Table 2: Organism identified based on sequence similarity of V3 region.
From the above, it is evident that the sequence of V3 region is a better indicator of bacterial diversity and truly reflected the rumen bacterial community. PCR-SSCP can bring down the cost and time required for sequence-based phylogenetic analysis. Therefore, it is recommended that the PCR-SSCP of V3 region as an alternative to creating DNA fingerprints to study the bacterial diversity and its identification in the rumen in place of cost-intensive metagenomic approaches.
The authors declare that they have no conflict of interest.
Download Provisional PDF Here
Article Type: RESEARCH ARTICLE
Citation: Kiruthiga R, Thanislass J, Antony PX, Lydia DC, Uma Maheswari D, et al. (2018) SSCP Analysis of V3 Region of 16S rRNA for the Characterization of Rumen Microbiome of Goat. J Anim Sci Res 2(1): dx.doi.org/10.16966/2576-6457.110
Copyright: © 2018 Kiruthiga R, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Publication history:
All Sci Forschen Journals are Open Access