Summary
Salmonella and Campylobacter are pathogens of concern frequently associated with poultry products. Production management intervention in breeder flocks and their progeny has potential to reduce the transmission of these bacteria. However, evidence of direct vertical transmission of these organisms through fertile turkey eggs to the progeny has not been demonstrated. The objective of this study was to determine if Campylobacter or Salmonella was transmitted through eggs and progeny of turkeys after artificial inoculation of breeder hens with semen containing marker strains of both bacteria (inoculated hens), as compared to uninoculated control hens. Eggs were collected and eggshells and yolks were checked for the presence of marker strains of Salmonella and Campylobacter. Eggs from both groups of hens were also set for incubation and hatch. At day of hatch, the number of hatched and unhatched eggs was recorded. Swabs from the hatchery trays containing eggshells and fecal residues were obtained, and paper pads with fecal droppings from both groups of poults, from inoculated and control hens, were also analyzed. Reproductive tract (RT) segments, including ovary and ovarian follicles, and upper level of the tract (from infundibulum to isthmus), of hens from both breeder flocks were analyzed at week 65. Semen and testes from toms were also analyzed. No evidence was obtained for vertical transmission of the marker strains through artificial insemination. However, Salmonella and Campylobacter were found in RT of hens and semen from toms, indicating a potential route of transmission of these foodborne pathogens to the progeny.
Description of Problem
Salmonella and Campylobacter are leading causes of bacterial gastroenteritis worldwide and both are frequently commensal colonizers of the intestinal tract of poultry. They have also been isolated from the avian reproductive tract and are, therefore, a possible source of contamination of hatching eggs and subsequent transmission to the progeny [1-5]. Furthermore, Salmonella and Campylobacter spp. in semen may result in possible transmission to the reproductive tract of hens via insemination [4,6,7]. Even though the outer and inner shell membranes of eggs offer protection against bacterial penetration, rapid penetration of Salmonella into the egg through pores or cracks in the shell has been demonstrated [8-11]. Sanitation of hatching eggs will not offer any protection to the developing embryo if bacteria have penetrated the eggshell membranes and into the internal contents [12]. Furthermore, egg incubation temperatures may enhance the multiplication of Salmonella. Although Salmonella is not found in high frequencies in eggs yolks in naturally contaminated eggs, the nutrient environment in the yolk is favorable for bacterial growth [9]. The vertical transmission of Campylobacter, through ovaries or oviduct to the egg, remains controversial. Although vertical transmission has been considered as a potential source of contamination to the progeny by some authors [4,13,14], it has been reported by others that transmission of Campylobacter through the eggs is a rare and unlikely event [15-17]. Nevertheless, these studies have been done primarily with chickens and there is a dearth of information concerning the mechanism of transmission of these pathogens in turkey hatching eggs. The present study was part of a research project where a turkey breeder flock was monitored from day of hatch to week 65 of life and evaluated for transmission routes of Salmonella and Campylobacter [18]. The current study was performed during the reproductive cycle of the breeder turkeys, week 32 to 65. The objectives of this research were to evaluate the potential for vertical transmission of both pathogens to eggs and progeny, and to assess any adverse effects of the presence of these bacteria on egg fertility and hatchability.
Materials and Method
Artificial Insemination and Egg Analysis
A nalidixic acid-resistant strain of Salmonella enterica serovar Enteritidis (S. Enteritidis) and a gentamicin-resistant strain of Campylobacter coli [19] were used to inoculate 80 breeder hens at week 39 via semen through artificial insemination. All bird handling and management procedures were approved by the NC State University Institutional Animal Care and use Committee. These inoculated hens (IH) were orally gavaged with the same marker strains at the beginning of the study, at day 10 and week 12 as described by Crespo, et al. [18]. A 0.5 mL aliquot of each bacterial culture, S. Enteritidis (approx. 108 cfu/mL) and C. coli (approx. 107 cfu/mL) were added to 2 mL of semen pooled from several inoculated toms (IT). IT was orally inoculated at the beginning of this study with marker strains of S. Typhimurium and C. jejuni [18]. A control flock of 20 hens, located in the opposite side of the same turkey house, was also inseminated using non-inoculated semen from the control toms (CT) that was diluted with Minnesota Turkey Growers Association (MTGA) semen extender containing gentamicin [20], added in the same ratio 1:2 (v/v) as the inoculated semen. Prior to the insemination of IH with semen containing marker strains of Salmonella and Campylobacter, both groups of hens were inseminated twice (week 35). The IH were artificially inseminated a total of four times using the semen containing both marker strains, at week 39, 40, 42, and 43. Insemination was performed with insemination styrene tubes of 10.3 cm in length and 0.208 cm of interior diameter [21], filled with approximately 0.035 mL of semen. Prior to the beginning of inseminations using the bacteria, six unwashed eggs from CH and 20 unwashed eggs from IH were analyzed. Eggs were collected every day, separated by pen number and kept in a cooler (12.8℃) until analysis. Eggs were analyzed in sets of six eggs from CH (unwashed). Five sets of unwashed eggs from IH were analyzed. The size of each set was 20 eggs except for one set of 10 due to unavailability of eggs (Table 1). Additionally, three sets of washed eggs from IH were tested. One of these sets with 20 eggs and two sets of 16 eggs due to unavailability of eggs (Table 1). A total of 24 eggs from CH and 142 eggs from IH were analyzed (90 unwashed eggs and 52 eggs sanitized after collection with a quaternary ammonium product following manufacturer’s dosage instructions [22]) (Table 1).
|
Hen Group |
Pre-insemination with bacteria |
Post-insemination with bacteria |
Total Salmonella- positive eggshells (%) |
| Set 1 |
Set 1 |
Set 2 |
Set 3 |
Set 4 |
Set 5 |
Post-insemination |
| Control (unwashed) |
6 |
6 |
6 |
6 (2)* |
0 |
6 |
2/24 (8) |
| Inseminated (unwashed) |
20 |
20 |
20 (2)* |
20 (3)* |
10 |
20 |
5/90 (6) |
| Inseminated (washed) |
0 |
0 |
0 |
16 |
20 |
16 |
0/52 (0) |
Table 1: Eggs analyzed before and after insemination of IH with semen containing marker strains of Salmonella and Campylobacter.
*Values in parenthesis indicate number of eggs (eggshells) positive for marker Salmonella.
A shell crush method as described by Musgrove, et al. [23] was used for bacterial recovery from the eggshells and yolks [23]. Eggs were cracked on the edge of a sterile beaker, egg whites were discarded and egg yolks pooled in a sterile beaker. The interior of the eggshell was rinsed with 20 mL of phosphate-buffered saline solution (PBS) [24] to eliminate residual egg whites. Eggshell and adhering membranes were crushed for one minute using a sterile glass rod in a sterile 50 mL centrifuge tube containing 20 mL of PBS. The sample was then divided in two sub-samples of 10 mL each for investigation of both pathogens. Buffered peptone water 2% (BPW) [25] was added for pre enrichment of Salmonella, and 2X Bolton broth (BB) (with supplements and laked horse blood added) [25] for the selective enrichment of Campylobacter. The crushed shells and membranes were also distributed in the two sub-samples. The pooled yolks were initially diluted with PBS (1:1 v/v), homogenized and divided in two sub-samples. BPW samples were incubated at 37℃ for 24 h, and BB samples were incubated at 42℃ for 24-48 h under microaerobic conditions (5% O2, 10% CO2, 85% N), using zip top bags filled with the gas mixture [26]. Brilliant green sulfa agar plates (BGS) [27] supplemented with 200 ppm of nalidixic acid [24] were used for recovering Salmonella and modified cefoperazone charcoal deoxycholate agar (mCCDA) [25] plates with 200 ppm gentamicin for the selective detection of Campylobacter. Plates were incubated at 37℃ aerobically for 24 h and 42℃ for 48 h under microaerobic conditions, respectively. Presumptive marker Salmonella colonies were confirmed by serology with Salmonella Poly O [28].
Hatch of Fertile Eggs
A total of 867 eggs, 675 from the IH flock and 192 from the CH flock (Set 1), were set in two different Jamesway incubators [29] at 37.5℃ and approximately 50% humidity. Eight days later, a second hatch was set with 450 eggs, 351 from IH and 99 from CH (Set 2). Transfer of the eggs was performed at day 24, and they remained in the hatchers until hatch on day 28. For both incubation groups, two different Jamesway hatchers were also used, both set at 36.9℃, with approximately 72% humidity. Hatchery trays containing eggshell residues and feces were swabbed at day of hatch, and pads with fecal droppings were also collected for analysis of Salmonella and Campylobacter. Eggs that did not hatch were identified, and the day of death was estimated based on the degree of development of the embryo. Cracked, rotten, and infertile eggs were also removed and recorded.
Analysis of Semen and Testes of Toms and Reproductive Tract of Hens
Semen was collected from both groups of toms at week 32 (sampled 2 different days) and week 33. Pooled semen from eight CT, and five and six IT, respectively, was collected by abdominal massage and analyzed for the presence of both bacteria. Three samples were analyzed for CT and six samples for IT. Enumeration of one sample per group (CT and IT) was performed in one occasion. For enumeration, 0.1 mL of semen was combined with 0.9 mL of buffered peptone water (BPW) and homogenized, serial dilutions were plated (0.1 mL) onto mCCDA. The detection limit was 100 cfu/mL. For Campylobacter detection, a 0.1 mL aliquot of semen was directly spread plated onto mCCDA and incubated 48 h at 42℃ in microaerobic conditions. Another 0.1 mL was added into 6 mL of BB (with supplements and laked horse blood) for enrichment. After incubation in microaerobic conditions at 42℃ for 48 h, samples were streaked onto mCCDA. For Salmonella detection, 1 mL of semen was added into 6 mL of BPW for enrichment. The BPW samples were incubated at 37℃ for 24 h followed by a selective enrichment step, with 0.5 mL of the previously enriched solution added into Tetrathionate (TT) broth base and 0.1 mL into Rappaport-Vassiliadis (RV) broth [30]. Tubes were vortexed and incubated at 42℃ for 24 h. After incubation a loopful from each of the tubes was streaked onto BGS agar. Presumptive Salmonella colonies were stabbed and streaked in triple sugar iron agar and lysine iron agar slants. Slants were incubated for 24 h at 37℃. Isolates giving typical Salmonella reactions were streaked onto nutrient agar (NA) plates [31] for purification, and confirmed by serology with Salmonella Poly O [28] and Poly H antiserum [25]. Confirmed Salmonella isolates were additionally streaked onto BGS supplemented with 200 ppm of nalidixic acid for confirmation of marker strains.
Birds were periodically euthanized and carcass exteriors were sprayed with 70% ethanol prior to opening the abdominal cavity. Guidelines of the approved Institutional Use and Animal Care Committee’s protocol at North Carolina State University were followed. Testes were aseptically removed at week 33 and 38 and analyzed for the presence of both bacteria. Testes were weighed and macerated with a rubber mallet and BB (with supplements and laked horse blood added) or BPW were added at a ratio of three times weight to volume of each sample for the enrichment of Campylobacter and Salmonella, respectively. Additional steps were the same as described for semen analysis. Reproductive tract of breeder hens was collected at week 65 and ovarian follicles and the upper segment of the tract (infundibulum, isthmus and magnum) were analyzed. Each sample was divided in two for analysis of each pathogen. Samples were processed as described for testes.
Progeny Monitoring
At day of hatch, 496 poults from inoculated hens (IP) and 126 poults from control hens (CP), were placed in 24 and 7 pens, respectively. Pens were located in opposite sides of a grow out house. Fecal samples from both groups were analyzed weekly to determine the presence of both bacteria. Pooled fecal droppings from the different pens from each group of poults were directly streaked onto mCCDA for detection of Campylobacter. Incubation conditions were the same as in previous sections. For Salmonella identification, feces were diluted in BPW (1:10) and subsequent steps were as described in the previous section.
At week one and five, 20 poults were euthanized. Organs (spleen, liver, and gallbladder) and gastrointestinal tracts were removed for analysis. Due to the small size of the poults, organs and gastrointestinal tracts were pooled as a composite of viscera, and treated as one sample. Samples were processed the same as described for testes. Feed, wood shavings and darkling beetle larvae collected in the progeny house were also analyzed (Table 2).
| Sampling Date |
Sample (quantity) |
Salmonella |
Campylobacter |
| Feb13 (week 1) |
Feed (25g), wood shavings (25g), darkling beetle larvae (25-30) |
Not Detected |
Not Detected |
| Mar13 (week 5) |
Darkling beetle larvae IP (25-30) |
Not Detected |
Not Detected |
| Mar13 (week 5) |
Darkling beetle larvae CP (25-30) |
Positive |
Not Detected |
Table 2: Vectors and other environmental samples analyzed.
IP: poults from inoculated hens; CP: poults from control hens.
Campylobacter and Salmonella Preservation
Campylobacter colonies were sub-cultured on MuellerHinton agar (MHA) [25] for purification, and nutrient agar was used for Salmonella. Isolates were preserved at -80℃ in cryovials containing brain heart infusion broth (BHI) with 20% glycerol.
Campylobacter and Salmonella Subtyping
Species of Campylobacter isolates were determined by multiplex polymerase chain reaction (PCR) using primers for amplifying the C. jejuni specific hipO gene (hipF 5’-ATG ATG GCT TCT TCG GAT AG-3’ and hipR 5’-GCT CCT ATG CTT ACA ACT GC-3’), and the C. coli ceuE gene (ceuF 5’-ATG AAA AAA TCT TTA GTT TTT GCA-3’ and ceuR 5’-GAT TTT ATT ATT TGT AGC AGC G-3’) [32-34]. Salmonella isolates were serotyped by SMART, a multiplex PCR and capillary electrophoresis analysis [35]. Campylobacter isolates were analyzed by pulsed-field gel electrophoresis (PFGE) after DNA restriction with SmaI [36]. Genetic relatedness based on the on the PFGE banding patterns were analyzed using BioNumerics [37]. Dice coefficient and unweighted pair group method with arithmetic mean (UPGMA), with 1.5% optimization and 1.7% position tolerance were used.
Campylobacter Antibiotic Resistance Determinations
Campylobacter isolates were tested for resistance to a panel of antibiotics (tetracycline, streptomycin, erythromycin, kanamycin, nalidixic acid, ciprofloxacin and gentamicin) based on their growth in the presence of specific amounts of the indicated antibiotic. Antibiotics and concentrations tested included kanamycin (25 µg/mL), nalidixic acid (20 µg/mL), ciprofloxacin (4 µg/mL), gentamicin (200 µg/mL), all four were obtained from Fisher [24], and tetracycline (16 µg/mL), erythromycin (10 µg/mL) and streptomycin (15 µg/mL) from Sigma [38]. All isolates were simultaneously also spotted on MHA to ensure viability. The C. jejuni ATCC 33560 (American Type Culture Collection; sensitive to all tested antibiotics) was included each time as quality control strain.
Statistical Analysis
Two-sided Fisher’s exact test was used to test independency between egg treatments. Fisher’s test was performed using JMP 11 software [39]. In hatch experiments data sets were analyzed using the GLM procedure of SAS [40]. Significance of data was set at P ≤ 0.05 in all cases.
Results and Discussion
Artificial Insemination and Egg Analysis
Salmonella and Campylobacter were not detected in the first set of eggs analyzed prior to insemination using the inoculated semen. Eggs analyzed from hens inseminated with semen containing marker strains of Salmonella (S. Enteritidis NAL) and Campylobacter (C. coli GK), were positive for S. Enteritidis NAL in five (6%) of the 90 unwashed eggshells analyzed; in turn, S. Enteritidis NAL was not found in eggshells of washed eggs (Table 1). In eggs from CH, S. Enteritidis NAL (marker strain inoculated into IH through semen) was isolated in two (8%) of the 24 eggshells analyzed (Table 1). The isolation of the marker Salmonella strain from control eggs was likely due to cross-contamination during egg collection or handling since CH and IH were placed in the same turkey house, albeit in different areas. Eggs were collected daily, and placed in plastic racks in a cooler, separating the eggs from CH and IH. The eggs were then transported from the farm to the laboratory for analysis. Previously published findings suggest that horizontal transmission via rodents or insects was possible [19]. Campylobacter was not detected in any of the eggs tested, either eggshells or yolks. The fact that eggs were held in a cooler at 12.8℃ could have a negative impact in Campylobacter survival in eggshells, leading to inability of detection by culture [16]. Furthermore, S. Enteritidis NAL was not detected in any of the yolks analyzed.
Based on two-sided Fisher’s exact test, there were no significant differences (P ≤ 0.05) between the eggs from either IH or CH. The small number of eggs analyzed was a limitation of the study, leading to low statistical power. Therefore, considering the low frequency of transmission of Salmonella through eggs in Salmonella-positive hens [9,12], the number of eggs and egg contents tested should be increased in future studies. In addition, the fact that Salmonella was not detected in any of the washed eggs, suggests a positive effect of washing and sanitizing the eggs on decreasing the potential of Salmonella transmitted through the eggshell.
Hatch Experiments
Eggs from CH hatched at 72.5% from first hatch (n=190), and 75% in the second hatch (n=98). Eggs from IH hatched at 78.1% rate for the first set (n=675), and 80.7% for the second set (n=351) (Table 3). Significant differences in fertility were found between treatments (Control, Inoculated) (p=0.0023), with higher fertility of in eggs from IH, but not between trials (Set 1, Set 2) (Table 3). The number of eggs used for the experiments was lower than typically employed under commercial conditions, which may affect the results. Also, the differences in numbers of eggs used per group of hens may have an effect. This limitation should be addressed in future experiments, via setting a higher number of eggs to get a better representation of commercial conditions, as well as similar numbers of eggs from both groups of hens (CH, IH). Hatchability rates were significantly different between treatments (Control/ Inoculated) (p= 0.0056) with a higher hatching rate on eggs from the IH. However, differences in hatchability between groups might be due to slight differences in temperature and humidity in the incubators used. There was no significant difference between the two sets of eggs from IH. Salmonella and Campylobacter were not detected in hatchery swabs with eggshell residues and fecal material, or on paper pads with fecal droppings analyzed at day of hatch from both groups (CH and IH).
| Set |
Parent flock |
Total eggs |
Infertile eggs (%) |
Poults hatched |
% Hatched
(total eggs /fertile eggs) |
Death rate first 7
days of incubation |
% Internal pip (26 days) |
% External pip
(27 days) |
| 1 |
Control |
190 |
19 (10%) |
124 |
65.3/72.5a |
0.09 |
1.58 |
9.47 |
| 1 |
Inoculated |
675 |
40 (5.9) |
496 |
73.5/78.1b |
0.08 |
0.89 |
8.74 |
| 2 |
Control |
98 |
14 (14.3) |
63 |
64.3/75.0a |
0.07 |
0 |
12.24 |
| 2 |
Inoculated |
351 |
24 (6.8) |
264 |
75.2/80.7b |
0.09 |
0 |
5.41 |
Table 3: Hatch experiment. Poults hatched, early death rates and pips.
a-b Values without a common superscript letter are significantly different (P<0.05), as analyzed by two-sided Fisher’s exact test.
Semen, Testes and Reproductive Tract of Hens
Both species of Campylobacter, C. jejuni and C. coli, were identified in semen, but none of the isolates were the marker strains initially inoculated orally into the toms (C. jejuni TSKQ) or hens (C. coli GK) [18]. All samples analyzed (CT=3; IT=6), were positive for Campylobacter. Since samples were directly streaked onto mCCDA, and also enriched, multiple colonies were analyzed per sample. A total of eight isolates were recovered for CT, and 11 isolates for IT. From the 19 isolates tested, 11 (58%) were C.jejuni resistant to tetracycline, five (26%) were C. coli resistant to kanamycin, and the remaining three (16%) were pan-sensitive C. jejuni.
Distribution of the isolates by group is shown in Figure 1. Four Campylobacter isolates from CT (50%) and seven (64%) from IT were C. jejuni resistant to tetracycline. Three (38%) of the isolates from CT and two (18%) from IT were C. coli resistant to kanamycin. Lastly, one (13%) isolate from CT and two (18%) from IT were pan-sensitive C. jejuni (Figure 1). Enumeration of two semen samples indicated Campylobacter levels of 6.0 × 102 cfu/mL and 9.0 × 102 cfu/mL in CT and IT, respectively. Both kanamycin-resistant C. coli and tetracycline-resistant C. jejuni were previously detected in fecal samples from both groups of toms (CT, IT); however, pan-sensitive C. jejuni was not detected in feces from toms. Furthermore, S. Agona was identified in one of the semen sample from CT. S. Agona was also isolated from fecal samples, cecum and jejunum of toms during the same project [18]. However, neither Campylobacter nor Salmonella were detected in testes. This, plus the lack of an aseptic semen collection technique suggest the possibility that semen was contaminated via contact with the cloaca.
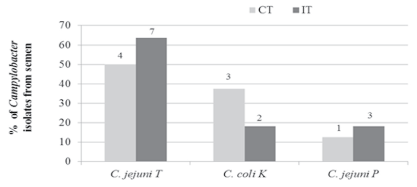
Figure 1: Percentage of Campylobacter species and antibiotic resistance profile isolated from semen samples. T- tetracycline; K-kanamycin; P-pansensitive. Frequencies were calculated based on total positive isolates per group (CT=8, IT=11). CT-Control Toms; IT-Inoculated Toms.
Analysis of reproductive tract samples from breeder hens yielded S. Enteritidis nalidixic acid-resistant (marker strain) and pan-sensitive C. jejuni from the upper segment of the reproductive tract (infundibulum, isthmus and magnum). However, neither of the bacteria was detected in ovaries or follicles. From the 20 hens analyzed from each group, pansensitive C. jejuni was detected in the upper segment of the reproductive tract in two IH and one CH Salmonella was isolated from three of the upper segments in IH, but not detected in CH. All three Salmonella isolates were the marker strain (S. Enteritidis NALR).
Progeny Monitoring
Feed and wood shavings analyzed were negative for both bacteria (Table 2). At week 1 (February 2013), S. Typhimurium (nalidixic acid sensitive) was isolated from fecal samples on CP, however it was not detected in IP until week 10 (April 2013) (Table 4). Furthermore, S. Typhimurium (nalidixic acid sensitive) was isolated from darkling beetle larvae (Tenebrionidae family) collected outside the pens of CP during week 5 (March 2013) (Table 2). Inspite of the presence of Salmonella in the environment and the presence of vectors for potential horizontal transmission, Salmonella was not detected in IP until week 10.
| |
Poults from CONTROL flock
(CP) |
Poults from INOCULATED
flock (IP) |
| Week |
Salmonella1 |
Campylobacter2 |
Salmonella1 |
Campylobacter2 |
| 1 |
+ |
- |
- |
- |
| 2 |
- |
- |
- |
- |
| 3 |
+ |
- |
- |
- |
| 5 |
- |
- |
- |
- |
| 6 |
+ |
- |
- |
- |
| 7 |
+ |
- |
- |
- |
| 8 |
+ |
- |
- |
- |
| 9 |
+ |
- |
- |
- |
| 10 |
ND3 |
- |
+ |
- |
| 11 |
+ |
- |
- |
- |
| 12 |
+ |
- |
- |
+ |
| 13 |
ND |
+ |
ND |
+ |
| 15 |
- |
+ |
- |
+ |
Table 4: Salmonella and Campylobacter detected in fecal samples from both groups of poults (progeny) (CP and IP).
1Salmonella detected did not share the same antibiotic resistance profile as marker strains used in the study for inoculating the breeder parents.
2 Campylobacter detected did not share the same antibiotic resistance profile as marker strains used in the study for inoculating the breeder parents.
3 ND- not determined.
Campylobacter was not detected until week 12 (last week of April 2013) in IP, and week 13 (first week of May 2013) in CP (Table 4). All Campylobacter isolates were pan-sensitive C. jejuni. Such findings suggest a more rapid spread of Campylobacter, and a similar pattern was also observed during monitoring of the breeder parents of these poults (data not shown). In previous studies it was reported that flocks were 100% colonized by day 7 to week 3 [41-43]. Colonization by C. jejuni at week 3 to 4 was thought to be related to the disappearance of maternal antibodies against Campylobacter in broiler chickens [44]. However, variation in colonization time, or even lack of colonization in turkey flocks have also been reported [42,45,46].
During monitoring of fecal samples of the breeder parents, naturally occurring strains of Campylobacter were detected at week 4 (second week of April) [18]. In both cases, other flocks were present in the houses prior to the placement of the poults (parent breeders and progeny). In the case of the parent breeders the house was empty for about three months prior to their placement. The house where the progeny were placed at day of hatch was empty about 4.5 months prior to the placement of the poults. Cleaning and sanitation in both houses was similar; with the exception that in the progeny house a disinfectant was not sprayed before placing the poults. Another difference could be the time of the year and temperature. Parent breeders were placed in the house during the second half of March 2012 (March 20th), while the progeny were placed at the beginning of February 2013 (February 5th). For both breeders and progeny, naturally occurring strains of Campylobacter were first detected in April. Environmental conditions such as temperature, humidity and litter moisture were not measured during the project. These parameters may impact the environmental spread of Campylobacter and Salmonella, and should be considered in subsequent studies. As previously reported for the breeder parents [18], Salmonella was detected early in CP (week 1) but was not detected in IP until 9 weeks later, suggesting a slower rate of spread. On the other hand, Campylobacter was detected later in fecal samples (week 12 and 13). It was first detected in IP at week 12, and then detected during the next sampling in the control group (CP) at week 13, demonstrating a faster ability to spread within the house.
At day 8, only one of 10 viscera composites from the CP was positive for Salmonella (Table 5). However, Salmonella was not identified in IP. Campylobacter was not detected in viscera of any of the groups (CP, IP). At day 36, six of the 10 CP viscera composites were positive for Salmonella, showing an increase in the number of birds colonized in the same area of the house, even when they were in different pens. All samples from IP were negative (Table 5). Campylobacter was not detected in any of the groups. All the Salmonella isolates were characterized as S. Typhimurium (nalidixic acid sensitive) by multiplex PCR and capillary electrophoresis analysis [35]. As with Campylobacter, the marker Salmonella strain inoculated through the semen was not isolated in fecal samples or intestinal samples in the poults. Nevertheless, the relatively low number of birds used for this study could affect the ability of detecting an event that occurs infrequently.
| |
Control Parent Flock |
Inoculated Parent Flock |
| Poults age |
Campylobacter |
Salmonella |
Campylobacter |
Salmonella |
| 8 days |
0/10 |
1/101 |
0/10 |
0/10 |
| 36 days |
0/10 |
6/101 |
0/10 |
0/10 |
Table 5: Campylobacter and Salmonella isolated from viscera composites of poults at day 8 and day 36. (Number of positive samples/ number of birds analyzed).
Comparison of Campylobacter Isolates from Progeny Feces, Reproductive Tract of Hens, and Semen of Toms
PFGE was performed to determine whether the multiple pan-sensitive C. jejuni isolates were representatives of the same strain. The three pan-sensitive C. jejuni isolates from the upper segment of the reproductive tract of breeder hens, pan-sensitive C. jejuni isolates from fecal samples from the poults, and two pan-sensitive C. jejuni isolates from semen of CT and IT toms were tested. The PFGE profiles of the two isolates from IH reproductive tract (IH6 RT and IH7 RT) were undistinguishable (Figure 2). Furthermore, the same band pattern was observed in C. jejuni isolates from the reproductive tract of CH (CH4 RT), and fecal samples of CP (CP04) and IP (IP1 02) (Figure 2). The other group clustered C. jejuni pan-sensitive isolated from fecal samples of CP (CP03) and IP (IP05, IP06 and IP1 01), showing a band pattern similar to the one observed in reproductive tract of CH (CH4 RT) (Figure 2). Isolates from semen of breeder toms were different from the other pan-sensitive C. jejuni strains isolated from breeder hens (CH and IH) and from the progeny (CP and IP) (Figure 2). A hypothesis for this observation can be the possibility of different environmental origin or a different time of entrance in the flock. Breeder toms were located in the same house throughout the study, and semen samples were collected at week 32 and 33. However, the hens were moved to a different house for the lighting program prior to being inseminated, and they remained in this house for 12 weeks before returning to the initial breeder house, being exposed to new environments, and the collection and analysis of reproductive tract as performed at week 65. In addition, the progeny were initially placed in a different grow out house until week 14 and then moved to the breeder house where they remained until week 16. Since Campylobacter was detected several months later in fecal samples of the older poults (week 12 and 13 in IP and CP, respectively), it may be suggested that colonization of the second-generation progeny was likely due to horizontal, rather than vertical transmission. Another observation was that Campylobacter isolates from the reproductive tract of CH (CHC4 RT) and IH (IHC6 RT and IHC7 RT) had an undistinguishable band pattern with isolates from ceca from CH (CHC4 ceca) and IH (IHC6 ceca), respectively (Figure 3), which may be possibly due to retrograde contamination from the cloaca into the reproductive tract. Furthermore, the three different band patterns (A, B and C) observed for these pan-sensitive C. jejuni isolates (Figure 3), indicates diversity among these C. jejuni isolates, suggesting different environmental origins or times of entrance in the flock.

Figure 2: SmaI PFGE profiles of pan sensitive C. jejuni isolates from reproductive tract (RT) of hens, semen of toms (CT and IT), and fecal samples from both groups of the progeny (CP and IP). CH-control hens; IH-inoculated hens; RT-reproductive tract; IT-inoculated toms; CT-control toms; CP-poults from control hens; IP-poults from inoculated hens. Letters and number indicate pen number and/or isolate number. The age of the turkeys at the moment of sampling is in parenthesis.
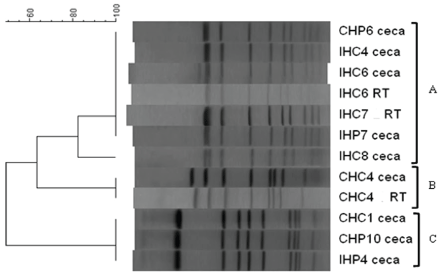
Figure 3: SmaI PFGE profiles of pan-sensitive C. jejuni isolates from the upper segment of reproductive tract (RT) and ceca of hens. CH-control hens; IH-inoculated hens; RT-reproductive tract. Letters and number indicate pen number. A,B and C, indicate different band patterns.
Based on the results of this study, the authors provide insights for future projects that can assist poultry researchers and the poultry industry in efforts to better understand the uncertainties regarding vertical transmission of Campylobacter, and reduce or eliminate Salmonella from turkey fertile hatching eggs and breeder flocks. A limitation in this study was the small number of samples analyzed (eggs, reproductive tract tissues). If the frequency of occurrence of vertical transmission is low, significantly larger numbers of animals, tissues, etc. will be required for detection of the pathogens in the progeny. Also, if the frequency of occurrence is extremely low, it may not be possible to ever demonstrate this with studies such as this one. If this is the case, more sensitive methods such as qPCR or sequencing-based methodologies could be used.
The fitness of the marker strains used for the study can also affect the outcome of colonization of reproductive tissues. During previously described monitoring of fecal and intestinal samples [18], naturally occurring strains seemed to outcompete marker strains and persist longer in the flock. In general, it would be important to replicate these experimental trials and compare the results.
Conclusions and Applications
- The presence of Salmonella and Campylobacter in semen during artificial insemination of the breeder hens did not adversely impact egg fertility and hatchability in this experiment.
- Based on the results of this study, washing and sanitizing hatching eggs may decrease the horizontal transmission of Salmonella through the eggshell.
- Artificial insemination of hens with semen contaminated with Salmonella or Campylobacter could not demonstrated vertical transmission of either bacteria to eggs or poults. Further study is needed to investigate this potential route of transmission.
- Better control of environmental (naturally occurring) strains of bacteria should be considered.
Acknowledgment
The present study was supported in part by the North Carolina Agricultural Foundation and the US Poultry and Egg Association (USPEA). Special thanks to Debbie Ort, Christina Shenton, Mike Mann, the farm crew, and lab members for assistance and technical support. Appreciation is also extended to Jonathan Frye and Lari Hiott (USDA-ARS-RRC, Athens, GA) for serotyping of Salmonella isolates.
